| "Evolution" mixes two
things together, one real, one imaginary.
Variation (microevolution) is
the real part. The types of bird beaks, the colors of
moths, leg sizes, etc. are variation.
Each type and length of beak a finch can have is already in the gene
pool and
adaptive mechanisms of
finches. Creationists have always agreed that there is variation within
species. What evolutionists do not want you to know is
that there are strict limits to variation that are never crossed,
something every breeder of animals or plants is aware of. Whenever
variation is pushed to extremes by selective breeding (to get the
most milk from cows, sugar from beets, bristles on fruit flies,
or any other characteristic), the line becomes sterile and dies
out. And as one characteristic increases, others diminish.
But evolutionists want
you to believe that changes continue, merging gradually into new kinds of
creatures. This is where the imaginary part of the theory of evolution
comes in.
It says that new information is added to the gene pool by mutation and
natural selection to create frogs from fish, reptiles from frogs, and
mammals from reptiles, to name a few. Do these big changes (macroevolution) really happen? Evolutionists tell us we cannot see evolution taking place because it happens too slowly. A human generation takes about 20 years from birth to parenthood. They say it took tens of thousands of generations to form man from a common ancestor with the ape, from populations of only hundreds or thousands. We do not have these problems with bacteria. A new generation of bacteria grows in as short as 12 minutes or up to 24 hours or more, depending on the type of bacteria and the environment, but typically 20 minutes to a few hours. There are more bacteria in the world than there are grains of sand on all of the beaches of the world (and many grains of sand are covered with bacteria). They exist in just about any environment: hot, cold, dry, wet, high pressure, low pressure, small groups, large colonies, isolated, much food, little food, much oxygen, no oxygen, in toxic chemicals, etc. There is much variation in bacteria. There are many mutations (in fact, evolutionists say that smaller organisms have a faster mutation rate than larger ones17). But they never turn into anything new. They always remain bacteria. Fruit flies are much more complex than already complex single-cell bacteria. Scientists like to study them because a generation (from egg to adult) takes only 9 days. In the lab, fruit flies are studied under every conceivable condition. There is much variation in fruit flies. There are many mutations. But they never turn into anything new. They always remain fruit flies. Many years of study of countless generations of bacteria and fruit flies all over the world shows that evolution is not happening today. |
|
|
|
Here is how the imaginary part is supposed to happen: On rare occasions a mutation in DNA improves a creature's ability to survive, so it is more likely to reproduce (natural selection). That is evolution's only tool for making new creatures. It might even work if it took just one gene to make and control one part. But parts of living creatures are constructed of intricate components with connections that all need to be in place for the thing to work, controlled by many genes that have to act in the proper sequence. Natural selection would not choose parts that did not have all their components existing, in place, connected, and regulated because the parts would not work. Thus all the right mutations (and none of the destructive ones) must happen at the same time by pure chance. That is physically impossible. To illustrate just how hopeless it is, imagine this: on the ground are all the materials needed to build a house (nails, boards, shingles, windows, etc.). We tie a hammer to the wagging tail of a dog and let him wander about the work site for as long as you please, even millions of years. The swinging hammer on the dog is as likely to build a house as mutation-natural selection is to make a single new working part in an animal, let alone a new creature.

Believing in beneficial mutations is like believing a short-circuit in the motherboard of your computer could improve its performance. To make any lasting change, a beneficial mutation would have to spread ("sweep") through a population and stay (become "fixed"). To evolutionists, this idea has been essential for so long that it is called a "classic sweep", "in which a new, strongly beneficial mutation increases in frequency to fixation in the population." Some evolutionist researchers went looking for classic sweeps in humans, and reported their findings in the journal Science. "To evaluate the importance of classic sweeps in shaping human diversity, we analyzed resequencing data for 179 human genomes from four populations". "In humans, the effects of sweeps are expected to persist for approximately 10,000 generations or about 250,000 years." Evolutionists had identified "more than 2000 genes as potential targets of positive selection in the human genome", and they expected that "diversity patterns in about 10% of the human genome have been affected by linkage to recent sweeps." So what did they find? "In contrast to expectation," their test detected nothing, but they could not quite bring themselves to say it. They said there was a "paucity of classic sweeps revealed by our findings". Sweeps "were too infrequent within the past 250,000 years to have had discernible effects on genomic diversity." "Classic sweeps were not a dominant mode of human adaptation over the past 250,000 years." --Hernandez, Ryan D., Joanna L. Kelley, Eyal Elyashiv, S. Cord Melton, Adam Auton, Gilean McVean, 1000 Genomes Project, Guy Sella, Molly Przeworski. 18 February 2011. Classic Selective Sweeps Were Rare in Recent Human Evolution. Science, Vol. 331, no. 6019, pp. 920-924.
A 35-year experiment by evolutionists shows how things really work. Instead of waiting for natural selection, researchers forced selection on hundreds of generations of fruit flies. They used variation to breed fruit flies that develop from egg to adult 20% faster than normal. But, as usual when breeding plants and animals, there was a down side. In this case the fruit flies weighed less, lived shorter lives, and were less resistant to starvation. There were many mutations, but none caught on, and the experiment ran into the limits of variation. They wrote that "forward experimental evolution can often be completely reversed with these populations". "Despite decades of sustained selection in relatively small, sexually reproducing laboratory populations, selection did not lead to the fixation of newly arising unconditionally advantageous alleles." "The probability of fixation in wild populations should be even lower than its likelihood in these experiments." --Burke, Molly K., Joseph P. Dunham, Parvin Shahrestani, Kevin R. Thornton, Michael R. Rose, Anthony D. Long. 30 September 2010. Genome-wide analysis of a long-term evolution experiment with Drosophila. Nature, Vol. 467, pp. 587-590.
You may have heard of the famous Lenski experiment. Dr. Richard E. Lenski is an evolutionary biologist who began a long-term experiment on February 24, 1988 that continues today. It looks for genetic changes in 12 initially identical populations of Escherichia coli bacteria that have been adapting to conditions in their flasks for over 60,000 generations. I have simplified a report by Scott Whynot, who studied 26 peer-reviewed scientific articles authored by Dr. Lenski (with others) published between 1991 and 2012. These papers represent the major genetic findings from 21 years of the experiment.
1. There was an insertion mutation that inhibited transcription of DNA involved in cell wall synthesis.
2. There was an insertion mutation in a regulatory region that encodes two proteins involved with cell wall synthesis. This may have led to larger cells.
3. A mutation in a gene led to a defect in DNA repair.
4. An insertion mutation may have knocked out a gene involved in programmed cell death and response to stress.
5. There was another mutation in a gene involved in response to stress, disrupting its function.
6. There was a mutation in the gene that encodes an enzyme that loosens DNA coils, leading to an increase in DNA supercoiling.
7. There was an insertion mutation in a gene that represses the production of nicotinamide adenine dinucleotide (NAD), a molecule that participates in many metabolic reactions, some affecting longevity. This might allow more NAD production.
8. The researchers noted an insertion mutation that they think inactivated a gene, resulting in greater glucose uptake. Glucose is a limited energy source in the experiment.
9. Deletion mutations caused the loss of the ability to catabolize D-ribose, an energy source that is not available in the experiment.
10. There was a mutation in a gene regulating transport of the sugar maltose, an energy source that is not present in the experiment.
11. The E. coli began to utilize an energy source, citrate, that they normally could not use in the presence of oxygen. E. coli already have the ability to transport and metabolize citrate where there is no oxygen, but they do not produce an appropriate transport protein for an environment with oxygen. In E. coli DNA, the gene for the citrate transporter that works without oxygen is directly upstream from genes for proteins with promoters that are active in the presence of oxygen. A replication of the region happened to put the transporter gene next to one of these promoters, so it could now be expressed in the presence of oxygen.
Except for number 11, the changes found in over 60,000 generations of bacteria were due to the disruption, degradation, or loss of genetic information. The ability to use citrate in the presence of oxygen, trumpeted by evolutionists as a big deal, was the result of previously existing information being rearranged, not the origin of new information. Mutations that result in a gain of novel information have not been observed.
Microevolution - Macroevolution
This candid admission is from the evolutionist journal Nature: "Darwin anticipated that microevolution would be a process of continuous and gradual change. The term macroevolution, by contrast, refers to the origin of new species and divisions of the taxonomic hierarchy above the species level, and also to the origin of complex adaptations, such as the vertebrate eye. Macroevolution posed a problem to Darwin because his principle of descent with modification predicts gradual transitions between small-scale adaptive changes in populations and these larger-scale phenomena, yet there is little evidence for such transitions in nature. Instead, the natural world is often characterized by gaps, or discontinuities. One type of gap relates to the existence of 'organs of extreme perfection', such as the eye, or morphological innovations, such as wings, both of which are found fully formed in present-day organisms without leaving evidence of how they evolved."-- Reznick, David N., Robert E. Ricklefs. 12 February 2009. Darwin's bridge between microevolution and macroevolution. Nature, Vol. 457, pp. 837-842.
Another evolutionary biologist wrote, "the processes underlying evolutionary innovation are remarkably poorly understood, which leaves us at a surprising conundrum: while biologists have made great progress over the past century and a half in understanding how existing traits diversify, we have made relatively little progress in understanding how novel traits come into being in the first place." "The origin of novel features continues to be a fascinating and challenging topic in evolutionary biology."-- Moczek, Armin P. May 2008. On the origins of novelty in development and evolution. BioEssays, Vol. 30, Issue 5, pp. 409-512.
Orphan genes - the final blow?
 |
|
Here
is an evolutionist with experience in molecular biology, Francois
Jacob. Francois Jacob won the Nobel Prize in Physiology or
Medicine in 1965, along with two others, for discoveries concerning
genetic control of enzyme and virus synthesis. He had joined
the Institut Pasteur in 1950. He was appointed Laboratory
Director there in 1956, then Head of the Department of Cell Genetics
in 1960. In 1964 he was appointed Professor at the College
de France, where a chair of Cell Genetics was created for him. He
was Chairman of the Board of the Institut Pasteur from 1982 to 1988.
The work of Francois Jacob dealt mainly with the genetic mechanisms
existing in bacteria and bacteriophages, and with the biochemical
effects of mutations. |
"During chemical evolution in prebiotic times and at the beginning of biological evolution, all those molecules of which every living being is built had to appear. But once life had started in the form of some primitive self-reproducing organism, further evolution had to proceed mainly through alterations of already existing compounds. New functions developed as new proteins appeared. But these were merely variations on previous themes. A sequence of a thousand nucleotides codes for a medium-sized protein. The probability that a functional protein would appear de novo by random association of amino acids is practically zero. In organisms as complex and integrated as those that were already living a long time ago, creation of entirely new nucleotide sequences could not be of any importance in the production of new information."20
For decades, everyone agreed. But as researchers compared the genes of similar creatures, they found that the genes differed, from just a little to a lot. They imagined different ways that could have happened. Gene duplication, non-deleterious frame shift mutations, alternative reading frames, overlap with transposable elements, horizontal gene transfer, or overlapping gene.45 As usual with evolutionists, they do not know what really happened, they assume it was one of these mental explanations, and that is enough. But some genes are so unique, even imagination fails. Evolutionists now conclude they must have assembled spontaneously - "de novo". In fact, "all genome and expressed sequence tag (EST) projects to date in every taxonomic group studied so far have uncovered a substantial fraction of genes that are without known homologs [equivalents]. These 'orphans' or 'taxonomically restricted genes' (TRGs) are defined as being exclusively restricted to a particular taxonomic group."21 "Orphan genes are defined as genes which lack detectable similarity to genes in other species". "They typically make up 10 to 30% of all genes in a genome."45
The foundation of evolution theory, gradual modification over time, slowly transforming genes that already exist, suddenly ran up against orphan genes, genes without parents in every taxonomic group studied so far. Looking at it objectively, the theory of evolution has been falsified. After careful study, evolutionists made a bold choice:
 __________________________________  |
They cut the theory's
last connection to reality, declaring that the impossible
is normal: of course genes are produced de novo! The
new foundation of evolution theory is Poof - there
it is (which sounds like the foundation of creation by Intelligent
Design - de novo). Evolutionists now think orphan genes are awesome. "There should be greater appreciation of the importance of the de novo origination of genes." "Today, we know that this evolutionary process is not impossible."47 "De novo evolution is clearly a strong force - constantly generating new genes over time." "It seems possible that most orphan genes have evolved through de novo evolution."35 "It looks as if we couldn't find the families of most orphans because they don't really have families."35 "The sequencing of a large number of eukaryotic and bacterial genomes has uncovered an abundance of genes without homologs... and has shown that new genes have arisen in the genomes of every group of organisms studied so far including humans".21 |
Origin of Life research
Evolutionists don't like to talk about "origin of life" research because it has been such a dead-end, but if chemicals never assembled themselves into the first living thing, evolution could never get started. So to keep hope alive, every once in a while over the last 60 years they have announced discoveries that supposedly bring us closer to understanding how life on Earth began. However, the main lesson scientists have learned over those decades is that the long molecules (polymers) that allow biological creatures to work must be isolated in pure concentrations for there to be any chance of success. Unfortunately, that can only happen in biochemistry labs, computer simulations, and living cells. In all other settings, the products are unusable due to contamination, unwanted reactions with other chemicals, and minuscule concentrations that quickly fall apart.
 Picture by T. Hoffman. |
|
Here is an origin-of-life
researcher, biochemist David
Deamer, who thought what he had made in the lab might
work in the real world. In 2005 he poured a concoction
of organic chemicals into a pool of hot water. He
was just trying to make the walls of a cell, like the
plastic case of a phone without the electronics inside.
Did it work? "The answer was a resounding
no." said another origin-of-life researcher. "The clays and metal ions present in the Siberian
pool blocked the chemical interactions." "Deamer's
demonstration that we cannot translate lab results to
natural settings is valuable." "This
provocative insight explains why the origin-of-life
field has been short on progress over the past half-century".
--Shapiro, Robert. 4 August 2011. Life's beginnings.
Nature, Vol. 476, pp. 30-31. |
Two prominent "origin-of-life" researchers have laid out their vision of how life arose from chemicals:
 |
|
1. Start with a molecule
capable of copying itself. "The first protocells
contained RNA (or something similar to it) and little else". 2. A fatty acid bubble forms around the self-copying molecule, which then makes a copy of itself with nucleotides that filter through the bubble. "Molecules as large as nucleotides can in fact easily slip across membranes as long as both nucleotides and membranes are simpler, more 'primitive' versions of their modern counterparts." |
4. "At some point some of the RNA sequences mutated, becoming ribozymes". The "ribozymes (folded RNA molecules analogous to protein-based enzymes) arise and take on such jobs as speeding up reproduction and strengthening the protocell's membrane. Consequently, protocells begin to reproduce on their own." "Other ribozymes catalyze metabolism -- chains of chemical reactions that enable protocells to tap into nutrients from the environment."
5. "Next, the organisms might have added protein-making to their bag of chemical tricks." "Complex systems of RNA catalysts begin to translate strings of RNA letters (genes) into chains of amino acids (proteins)." "Proteins take on a wide range of tasks within the cell."
6. "Protein-based catalysts, or enzymes, gradually replace most ribozymes." "Proteins would have then taken over RNA's role in assisting genetic copying and metabolism."
7. Later, the organisms would have 'learned' to make DNA". "Thanks to its superior stability, DNA takes on the role of primary genetic molecule. RNA's main role is now to act as a bridge between DNA and proteins."
8. "Organisms resembling modern bacteria adapt to living virtually everywhere on earth and rule unopposed for billions of years, until some of them begin to evolve into more complex organisms." --Ricardo, Alonso, Jack W. Szostak. September 2009. Life on Earth. Scientific American, pp. 54-61.
They are currently working on steps 1 and 2 in the laboratory.
Let's compare their "origin of life" ideas to the plans of children making a spaceship out of a cardboard box:
 |
|
1. Get a large box. Draw
controls and gauges on the inside. Cut out a door and round windows. Attach
cardboard fins to the sides. 2. Put a chair in the box, sit down and start the countdown. |
4. After passing Neptune, engage the warp drive in a direction perpendicular to the plane of the ecliptic to avoid the Kuiper belt.
The children are currently working on steps 1 and 2, and are as close to demonstrating their vision as the "origin-of-life" researchers are.
Gradual change versus leaps
There are two versions of evolution theory. The main version proposes that many tiny changes over millions of years made new creatures. It is called the Modern Synthesis or Neo-Darwinian evolution.
But "major transitions in biological evolution show the same pattern of sudden emergence of diverse forms at a new level of complexity." "The principal 'types' seem to appear rapidly and fully equipped with the signature features of the respective new level of biological organization. No intermediate 'grades' or intermediate forms between different types are detectable."23
Since the fossil record does not show long series of tiny changes between one type of creature and another, some evolutionists proposed a modification to evolution theory. It says that change occurred by occasional leaps (punctuated equilibrium), not gradually. However, each hypothetical beneficial mutation could only make a slight change. Any more than that would be so disruptive as to cause death. So punctuated equilibrium is not really about big leaps. It envisions a lot of slight changes over thousands of years, then nothing happens for millions of years. Evolutionists say with a straight face that no fossils have been found from a leap because thousands of years is too fast in the billions of years of "geologic time" to leave any. Yet without fossils there is no evidence that any leaps ever happened, and of course there is no evidence that leaps or gradual changes beyond variation are happening today in any of the millions of species that still exist.
Fossil record
Evolution is all about constant change, whether gradual or in leaps. Consider a cloud in the sky: it is constantly changing shape due to natural forces. It might look like, say, a rabbit now, and a few minutes later appear to be, say, a horse. In between, the whole mass is shifting about. In a few more minutes it may look like a bird. The problem for evolution is that we never see the shifting between shapes in the fossil record. All fossils are of complete animals and plants, not works in progress "under construction". That is why we can give each distinct plant or animal a name. If evolution's continuous morphing were really going on, every fossil would show change underway throughout the creature, with parts in various stages of completion. For every successful change there should be many more that lead to nothing. The whole process is random trial and error, without direction. So every plant and animal, living or fossil, should be covered inside and out with useless growths and have parts under construction. It is a grotesque image, and just what the theory of evolution really predicts. Even Charles Darwin had a glimpse of the problem in his day. He wrote in his book On the Origin of Species: "The number of intermediate varieties which have formerly existed on Earth must be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory." The more fossils that are found, the better sense we have of what lived in the past. Since Darwin's day, the number of fossils that have been collected has grown tremendously, so we now have a pretty accurate picture. The gradual morphing of one type of creature to another that evolution predicts is nowhere to be found. There should have been millions of transitional creatures if evolution were true. In the "tree of life" that evolutionists have dreamed up, gaps in the fossil record are especially huge between single-cell creatures, complex invertebrates (such as snails, jellyfish, trilobites, clams, and sponges), and what evolutionists claim were the first vertebrates, fish. In fact, there are no fossil ancestors at all for complex invertebrates or fish. That alone is fatal to the theory of evolution. The fossil record shows that evolution never happened.
What fossil evidence is there for the evolutionist vision for the origin of life? Nothing, except for shapes that might have been cells or something else. Everyone agrees that the big surprise is the sudden appearance of fossils above the bedrock in the Cambrian Explosion.
 |
|
The fossils of the
Cambrian Explosion are complex invertebrates, sea creatures
like trilobites, sponges, worms, jellyfish, sea urchins,
sea lilies, mollusks, brachiopods (lamp shells), sea
cucumbers, and swimming crustaceans such as |
 |
|
Opabinia, 3 inches
long (8 cm) with 5 eyes and a long claw arm, and |
 |
|
Anomalocaris, 3
feet long (91.5 cm), and the top predator in the
Cambrian environment. |
 |
|
In the classification
diagram biologists use, these animals are so unrelated
to each other that they are in different classes or
even phyla. From time to time evolutionists announce
with great fanfare that they have gotten a colony of
bacteria to eat something they could not eat before,
or some other small variation. These changes are
always below the family level on the diagram. If
evolution were true, there would have been ancestors
and transitional creatures between each genus, family,
order, class, and phylum in the layers below the Cambrian
Explosion. But there are no fossils
for any of these. |
 |
|
Fossil compound eyes
from the Lower Cambrian, where
the first complex creatures suddenly appear in the fossil
record, have been found in the
Emu Bay Shale of South Australia. The fossils
are supposedly about 515 million years old. They may be
corneas of Anomalocaris that were shed during moulting. The lenses
are packed tighter than Lower Cambrian trilobite eyes,
"which are often assumed to be the most powerful
visual organs of their time." Notice that
the lenses in the picture are different sizes. It
is the same in
the fossils. Each eye has "over 3,000 large ommatidial
lenses". "The arrangement and size-gradient
of lenses creates a distinct [forward] 'bright zone'...
where the visual field is sampled with higher light
sensitivity (due to larger ommatidia) and possibly higher
accuity". This indicates "that these
eyes belonged to an active predator that was capable
of seeing in low light." "The eyes are
more complex than those known from contemporaneous trilobites
and are as advanced as those of many living forms"
today, such as the fly in this picture, "revealing
that some of the earliest arthropods possessed highly
advanced compound eyes".27 When the earliest
form is the most complex, there is no evolution. |
 |
|
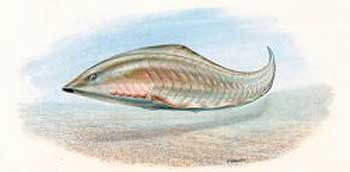
This tiny fish (a little
over an inch long, or 3 cm) is Haikouichthys. Its
fossils have also been found in the Lower Cambrian. This "first fish" has a spine
and spinal cord, eyes, gills, fins, scales, mouth,
etc., though no jaw, like a lamprey. About 500
were found buried together.39 |
 |
|
This is Guiyu, a fossil
fish that "represents the oldest near-complete
gnathostome (jawed vertebrate)."48
It measures about 15 inches long, or 37 cm. Clearly,
the earliest fish were as much fish as today's fish.
Guiyu is "a representative of modern fishes"
from the Silurian, before the so-called "age of
fishes" (Devonian).9
In the evolutionist's mind, "a whole series
of major branching events... must have taken place well
before the end of the Silurian." "A
significant part of early vertebrate evolution is unknown."9 |
 |
|
Coelacanth
disappeared
from the fossil record with the last of the
dinosaurs.
That was supposedly 65 million years ago.
In the early 1900s, evolutionists touted it as
the first walking fish, the transition between
fish
and tetrapods. That is, until 1938 when one was
found alive and unable to walk. Evolution
theory
says that pressures from competition and the
environment
force changes over time. In chapter
9 of his book, Darwin wrote of ancestor species
in general: "If, moreover, they had been the progenitors of these
orders, they would almost
certainly have been long ago supplanted and exterminated by their
numerous and
improved descendants." Here is a coelacanth
today, alive and unchanged like many "living
fossils". Where is the evolution? |
 |
|
Evolutionists
tell us this dragonfly has not shown up in the
fossil record for 250-300 million years! Dozens of the Ancient Greenling Damselfly live near Melbourne, Australia.
"The damselfly, part of the dragonfly
group Odonata, is the only living representative of the family Hemiphlebiidae.
Its ancient predecessors are found solely in 250-300 million-year-old fossil
records from Brazil to Russia." --Smith, Bridie.
January 5, 2010. Found: fossil-linked, listed damselfly.
www.theage.com.au (newspaper website)
|
 |
|
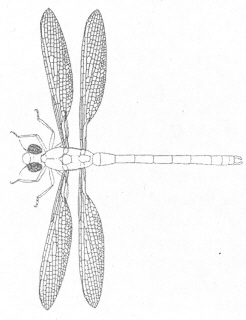
This is a drawing of a
supposed predecessor, Protozygoptera. With a wingspan
of under 6 cm, it is the
earliest damselfly-like insect ever found and "the
origin of modern dragonflies". Its fossil
wing was found in rocks of the
Upper Carboniferous which evolutionists think
are about 300 million years old. As with many creatures, dragonflies
appear suddenly in the fossil record, fully formed.
Damselflies living today look like Protozygoptera;
there are no transitional intermediates
and there was no evolution. --Jarzembowski, E.A.,
A. Nel. 2002. The earliest damselfly-like insect and
the origin of modern dragonflies (Insecta: Odonatoptera:
Protozygoptera). Proceedings of the Geologists' Association,
Vol. 113, pp. 165-169. |
 |
|
Evolutionists always
point to Archaeopteryx as the great example of a transitional
creature, appearing
to be
part dinosaur and part bird. However,
it is a fully formed, complete animal with no half-finished
components or useless growths. Most people know
"the stereotypical ideal of Archaeopteryx as a
physiologically modern bird with a long tail and teeth".
Research now "shows incontrovertibly that
these animals were very primitive". "Archaeopteryx
was simply a feathered and presumably volant [flying]
dinosaur. Theories regarding the subsequent steps
that led to the modern avian condition need to be reevaluated."
--Erickson, Gregory, et al. October 2009. Was
Dinosaurian Physiology Inherited by Birds? Reconciling
Slow Growth in Archaeopteryx. PLoS ONE, Vol.
4, Issue 10, e7390. |
What evolutionists now know for sure is that their celebrity superstar was not a transitional creature after all. Wow! OMG. They better find a new one fast...
 |
|
How about the Platypus?
They could call it a transitional creature between
ducks and mammals. The furry platypus has a
duck-like bill, swims with webbed feet, and lays eggs. |
 |
|
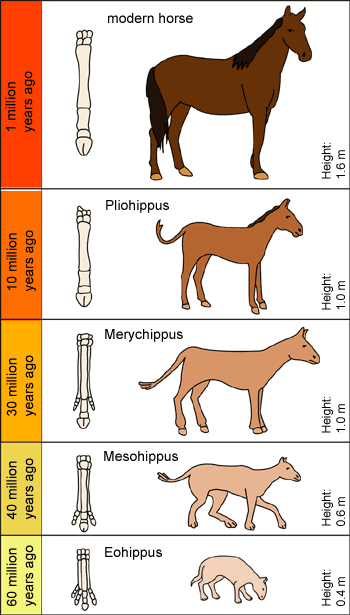
The same is true for
the famous horse series. Looks great, doesn't
it? But each of the supposed ancestors is
a complete animal. They
are not full of failed growths and there are no
parts under construction. There are many more
differences between each type of animal than their
size and the number of toes. Every change in structure,
function, and process would have had to develop
through random trial-and-error if evolution were true,
but no transitional forms have been found.
The fossils have not caught any
changes in the midst of being created, even though they
should
have occurred over long periods of time. In the
late 1800's, evolutionists
simply placed living and extinct species next to each
other to make the horse series. However, evolutionists
no longer believe there was the direct ancestry (orthogenesis)
shown in this chart... |
 |
|
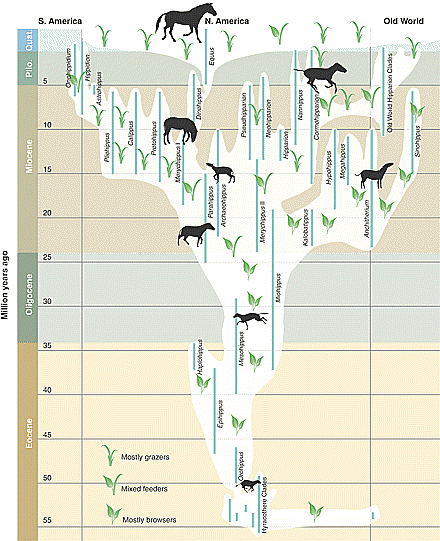
Evolutionists now imagine it to
be this branching bush. Many of the supposed ancestors
apparently lived at the same time, especially
after Mesohippus. It
is doubtful that Hyracotherium (formerly Eohippus) has
any connection to horses. So the progression
of toes is an illusion that was useful when the theory of evolution was first being sold
to the public. Several hundred species
are extinct; only one genus, Equus, survives. |
So the level of "evolution" in the horse
bush is within a family, which is pretty low and has no relevance
to the main issue, macroevolution. But evolutionists have
always used the varieties in the family Equidae to entice people
to imagine that all animals morph from one kind to another. Look
at their tree of life for mammals, the class Mammalia.
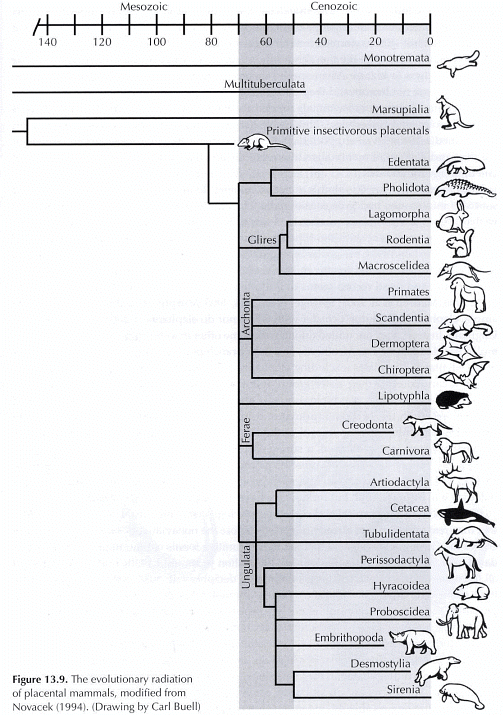
Novacek, M.J. 1994. The Radiation of Placental Mammals, in Prothero,
D.R. and Schoch, R.M. (editors) Major Features of Vertebrate
Evolution. Paleontological Society Short Courses in Paleontology,
No. 7, pp. 220-237.
That's
right; they want us to believe that elephants and manatees, primates
and tree shrews had common ancestors. They show you three
toes changing to one toe, browsing teeth changing to grazing teeth
in the horse bush so that you will believe that reindeer and whales
morphed from common ancestors. That is why the horse series
is an evolutionist icon.
New discoveries are bringing down the whole notion of a "tree of life", as passages from an article in the mainstream magazine New Scientist show:26 "The tree-of-life concept was absolutely central to Darwin's thinking, equal in importance to natural selection, according to biologist W. Ford Doolittle of Dalhousie University in Halifax, Nova Scotia, Canada. Without it the theory of evolution would never have happened." "For much of the past 150 years, biology has largely concerned itself with filling in the details of the tree. 'For a long time the holy grail was to build a tree of life,' says Eric Bapteste, an evolutionary biologist at the Pierre and Marie Curie University in Paris, France. A few years ago it looked as though the grail was within reach."
"But today the project lies in tatters, torn to pieces by an onslaught of negative evidence. Many biologists now argue that the tree concept is obsolete and needs to be discarded. 'We have no evidence at all that the tree of life is a reality,' says Bapteste. That bombshell has even persuaded some that our fundamental view of biology needs to change." "The problems began in the early 1990s when it became possible to sequence actual bacterial and archaeal genes". "As more and more genes were sequenced, it became clear that the patterns of relatedness could only be explained if bacteria and archaea were routinely swapping genetic material with other species - often across huge taxonomic distances". " 'There's promiscuous exchange of genetic information across diverse groups,' says Michael Rose, an evolutionary biologist at the University of California, Irvine." "As early as 1993, some were proposing that for bacteria and archaea the tree of life was more like a web. In 1999, Doolittle made the provocative claim that 'the history of life cannot properly be represented as a tree'.13 'The tree of life is not something that exists in nature, it's a way that humans classify nature,' he says."
"Recent research suggests that the evolution of animals and plants isn't exactly tree-like either." "A team at the University of Texas at Arlington found a peculiar chunk of DNA in the genomes of eight animals - the mouse, rat, bushbaby, little brown bat, tenrec, opossum, anole lizard and African clawed frog - but not in 25 others, including humans, elephants, chickens and fish. This patchy distribution suggests that the sequence must have entered each genome independently by horizontal transfer."34 "HGT [horizontal gene transfer] has been documented in insects, fish and plants, and a few years ago a piece of snake DNA was found in cows." "Biologist Michael Syvanen of the University of California, Davis... compared 2000 genes that are common to humans, frogs, sea squirts, sea urchins, fruit flies and nematodes. In theory, he should have been able to use the gene sequences to construct an evolutionary tree showing the relationships between the six animals. He failed."
"The problem was that different genes told contradictory evolutionary stories." " 'We've just annihilated the tree of life. It's not a tree any more, it's a different topology [design or shape] entirely,' says Syvanen." "It is clear that the Darwinian tree is no longer an adequate description of how evolution in general works." "Rose goes even further. 'The tree of life is politely buried, we all know that,' he says. 'What's less accepted is that our whole fundamental view of biology needs to change.' Biology is vastly more complex than we thought, he says." " 'The tree of life was useful,' says Bapteste. 'It helped us to understand that evolution was real. But now we know more about evolution, it's time to move on.' "26
Evolutionists write: "The meaning, role in biology, and support in evidence of the universal 'Tree of Life' (TOL) are currently in dispute. Some evolutionists believe... that we can with available data and methods reconstruct this tree quite accurately, and that we have in fact done so, at least for the major groups of organisms. Other evolutionists... do not doubt that some... branching tree can in principle represent the history of all life. Still other evolutionists, ourselves included, question even this most fundamental belief, that there is a single true tree." "Darwin claimed that a unique inclusively hierarchical pattern of relationships between all organisms based on their similarities and differences was a fact of nature." Yet "the only data sets from which we might construct a universal hierarchy including prokaryotes, the sequences of genes, often disagree and can seldom be proven to agree. Hierarchical structure can always be imposed on or extracted from such data sets by algorithms designed to do so, but at its base the universal TOL rests on an unproven assumption about pattern that, given what we know about process, is unlikely to be broadly true." There is "the possibility that hierarchy is imposed by us rather than already being there in the data."12 "The finding that, on average, only 0.1% to 1% of each genome fits the metaphor of a tree of life overwhelmingly supports the... argument that a single bifurcating tree is an insufficient model to describe the microbial evolutionary process." "When chemists or physicists find that a given null hypothesis can account for only 1% of their data, they immediately start searching for a better hypothesis. Not so with microbial evolution, it seems, which is rather worrying. Could it be that many biologists have their heart set on finding a tree of life, regardless of what the data actually say?"10 "A single, uninterrupted TOL does not exist, although the evolution of large divisions of life for extended time intervals can be adequately described by trees." "Tree topology tends to differ for different genes."26 The genomes of all life forms are collections of genes with diverse evolutionary histories." "The TOL concept must be substantially revised or abandoned because a single tree topology or even congruent topologies of trees for several highly conserved genes cannot possibly represent the history of all or even the majority of the genes." "The 'strong' TOL hypothesis, namely, the existence of a 'species tree' for the entire history of cellular life, is falsified by the results of comparative genomics." "So the TOL becomes a network, or perhaps most appropriately, the Forest of Life that consists of trees, bushes, thickets..., and of course, numerous dead trunks and branches."22
Kevin Peterson, a molecular paleobiologist at Dartmouth
College, "has been reshaping phylogenetic trees for the past few years, ever
since he pioneered a technique that uses short molecules called microRNAs to
work out evolutionary branchings. He has now sketched out a radically different
diagram for mammals: one that aligns humans more closely with elephants than
with rodents."
" 'I've looked at thousands of microRNAs, and I can't
find a single example that would support the traditional tree,' he says. The
technique "just changes everything about our understanding of mammal evolution'."
"And as he continues to look, he keeps uncovering problems,
from the base of the animal tree all the way up to its crown."
"Peterson and his team are now going back to mammalian
genomes to investigate why DNA and microRNAs give such different evolutionary
trajectories. 'What we know at this stage is that we do have a very serious
incongruence,' says Davide Pisani, a phylogeneticist at the National University
of Ireland in Maynooth". --Dolgin,
Elie. 28 June 2012. Rewriting Evolution. Nature, Vol.486, pp.460-462.
The big fudgeThere were warnings of problems in the very "tree of life" diagrams evolutionists were fabricating. If you believed these diagrams of new forms arising from old forms, then you believed that parts can be lost from a creature and then exactly re-invented in the same creature millions of years later. And that might happen several times! Not only that, the same part might appear independently in unrelated creatures. For example, wings would have had to evolve completely independently four times: in insects, flying reptiles, birds, and bats.
 |
|
When
a creature produces light with chemicals it is called
bioluminescence. "A remarkable diversity
of marine animals and microbes are able to produce
their own light, and in most of the volume of the
ocean, bioluminescence is the primary source of
light." "On land, fireflies are
the most conspicuous examples, but other luminous
taxa include other beetles, insects like flies and
springtails, fungi, centipedes and millipedes, a
snail, and earthworms." Evolutionists
think bioluminescence evolved independently 40 to
50 separate times!
--Haddock, Steven H.D., Mark A. Moline, James F.
Case. 2010. Bioluminescence in the Sea. Annual Review
of Marine Science, Vol. 2, pp. 443-493. |
Instead of heeding the warnings and scrapping the diagrams, they solved the problem by giving it a name: either "convergent" or "parallel" evolution (according to the situation), and it became standard in evolutionist writings. Now they just casually throw out statements like "convergent evolution is widespread across the mammal tree of life". --Helgen, Kristofer M. 28 October 2011. The Mammal Family Tree. Science, Vol. 334, pp. 458-459.
In their sales pitch to the public, evolutionists use the gimmick, "if a million monkeys typed randomly for millions of years, eventually one would type one of Shakespeare's sonnets", and people think "well, maybe...". How about the monkeys typing the same sonnet twice or more? It would be shocking if random mutation-natural selection produced any working part, let alone the same part twice or more from scratch. With convergent/parallel evolution, evolutionists could fudge any situation.
Perhaps the most brazen example of this can be seen in the "parallel evolution" of two types of mammals, placental (such as humans) and marsupial (such as kangaroos). Evolutionists tell us that each group evolved separately, yet many are remarkably similar, including cats, mice, wolves, moles, flying squirrels, anteaters, and others. This is whole animal duplication, not just an individual part.
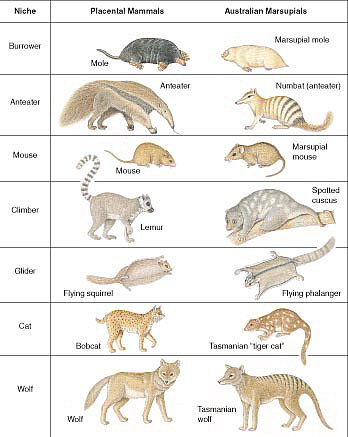
Design
flaws?  Evolutionists say that while no intelligent designer would
design anything with flaws, evolution is a mechanical process of trial and
error, so evolution easily explains the existence of flaws in biology. In his 1986 book, "The Blind Watchmaker," the
famous evolutionary biologist Richard Dawkins wrote about the eye, "Any engineer would
naturally assume that the photocells would point towards the light, with their
wires leading backwards towards the brain.
He would laugh at any suggestion that the photocells might point away,
from the light, with their wires departing on the side nearest the light. Yet this is exactly what happens in all
vertebrate retinas." "This means that
the light, instead of being granted an unrestricted passage to the photocells,
has to pass through a forest of connecting wires, presumably suffering at least
some attenuation and distortion".
Evolutionist professors have told this to their students for years,
while looking at them with perfect vision.
Evolutionists say that while no intelligent designer would
design anything with flaws, evolution is a mechanical process of trial and
error, so evolution easily explains the existence of flaws in biology. In his 1986 book, "The Blind Watchmaker," the
famous evolutionary biologist Richard Dawkins wrote about the eye, "Any engineer would
naturally assume that the photocells would point towards the light, with their
wires leading backwards towards the brain.
He would laugh at any suggestion that the photocells might point away,
from the light, with their wires departing on the side nearest the light. Yet this is exactly what happens in all
vertebrate retinas." "This means that
the light, instead of being granted an unrestricted passage to the photocells,
has to pass through a forest of connecting wires, presumably suffering at least
some attenuation and distortion".
Evolutionist professors have told this to their students for years,
while looking at them with perfect vision.
Evolutionists say that while no intelligent designer would
design anything with flaws, evolution is a mechanical process of trial and
error, so evolution easily explains the existence of flaws in biology. In his 1986 book, "The Blind Watchmaker," the
famous evolutionary biologist Richard Dawkins wrote about the eye, "Any engineer would
naturally assume that the photocells would point towards the light, with their
wires leading backwards towards the brain.
He would laugh at any suggestion that the photocells might point away,
from the light, with their wires departing on the side nearest the light. Yet this is exactly what happens in all
vertebrate retinas." "This means that
the light, instead of being granted an unrestricted passage to the photocells,
has to pass through a forest of connecting wires, presumably suffering at least
some attenuation and distortion".
Evolutionist professors have told this to their students for years,
while looking at them with perfect vision.
It is true that "images projected onto the retina have to
pass several layers of randomly oriented and irregularly shaped cells with
intrinsic scatterers before they reach the light-detecting photoreceptor cells".
"When light passes through multiple
layers of cells, as in tissues, images rapidly deteriorate".16
"The mammalian retina and the peripheral retina of humans
and primates are organized in a seemingly reverse order with respect to the
light path. This arrangement places the
photoreceptors, responsible for light absorption, as the last cells in the path
of light, rather than the first. Therefore,
the incident light must propagate through five reflecting and scattering layers
of cell bodies and neural processes before reaching the photoreceptors. This 'inverted' retinal structure is expected
to cause blurring of the image and reduction in the photon flux reaching the
photoreceptors, thus reducing their sensitivity."25
Then researchers took a closer look at glial cells in the
eye. "Glial cells are the most abundant
cell types in the central nervous system. They surround neurons to support and insulate them."
"Muller cells, the major type of glial cells
in the retina, are responsible for the homeostatic and metabolic support of
retinal neurons. By mediating
transcellular ion, water, and bicarbonate transport, Muller cells control the
composition of the extracellular space fluid. Muller cells provide trophic and
anti-oxidative support of photoreceptors and neurons and regulate the tightness
of the blood-retinal barrier. By the
uptake of glutamate, Muller cells are more directly involved in the regulation
of the synaptic activity in the inner retina. This review gives a survey of recently
discoved new functions of Muller cells. Muller cells are living optical fibers that
guide light through the inner retinal tissue."37
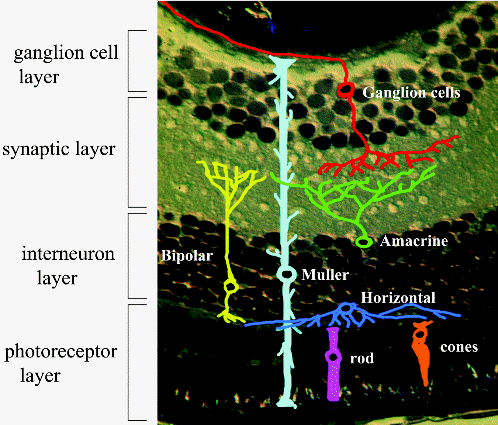
Muller cells span the entire thickness of the retina.
"Individual Muller cells act as optical
fibers. Furthermore, their parallel
array in the retina is reminiscent of fiberoptic plates used for low-distortion
image transfer. Thus, Muller cells seem to mediate image transfer through the
vertebrate retina with minimal distortion and low loss."16
"The basic fiberoptic plate-like structure is especially
characteristic for the retinae of all mammals with the exception of the fovea
centralis of humans and higher primates, the region of our retina that is
responsible for sharp vision; here, the photoreceptor cells are not obscured by
any inner retinal layers at all."16
"Every mammalian Muller cell is coupled to one cone
photoreceptor cell (responsible for sharp seeing under daylight conditions,
i.e., photopic vision) plus a species-specific number of rod photoreceptor
cells (about 10 in both man and guinea pig), serving low light level (scotopic)
vision."16
"Light of relevant wavelengths for cone visual pigments is
directed towards the cones, while light of wavelengths more suitable for rod
vision is allowed to leak outside the Muller cells towards the surrounding rods."25
"The fundamental
features of the array of glial cells are revealed as an optimal structure
designed for preserving the acuity of images in the human retina. It plays a crucial role in vision quality, in
humans and in other species."24
As usual, once
we learn more about something in biology, evolutionist claims fall
apart.
Reliving evolution?An old evolution myth still
hanging around is the notion that things that look like gill-slits, tails,
etc. in developing human embryos show the embryo repeating
all the stages of evolution. In 1866, Ernst Haeckel
proposed his "biogenetic law" (not to be confused with
the law of biogenesis that says life only comes from life). His
idea was that growing vertebrate embryos went through all the forms
of their supposed evolutionary ancestors ("ontogeny recapitulates
phylogeny"). He published drawings comparing growing
embryos of a number of animals such as the pig, cat, salamander,
etc. to growing human embryos. The similarities that he said
he found helped persuade people to believe the theory of evolution.
Scientists eventually discovered enough about embryology to
quietly discard the "biogenetic law", but it was not until
a careful photographic study of growing vertebrate embryos was conducted
in 1997 that Haeckel's deceit was fully revealed. They
found that his drawings were so far from reality that they could
not have been done from the actual embryos.38 He
must have faked them.Vanishing vestigialsIf evolutionists do not know what something does, they assume it is useless, as we will see with "junk DNA". One of their "proofs" of evolution has been that as creatures evolve, some body parts that were useful long ago become less important in the new and improved creatures. Eventually these parts no longer function and they shrink in size. Evolutionists called them "vestigial organs". In the late 1800s they made long lists of vestigial organs in humans, including the tonsils, pineal gland, thymus, and appendix.5 In the years since, advances in our understanding of anatomy and biology have knocked them off the lists one by one. Yet the notion lingers on that "there is something to it". In 2009, researchers at Arizona State and Duke Universities reported that the little appendix is a "safe house" for important gut bacteria. If the intestine becomes infected and is forced to flush everything out (diarrhea), the good bacteria stored in the appendix are there to return the intestine to normal working order.40,41 "Maybe it's time to correct the textbooks," says William Parker, Ph.D., assistant professor of surgical sciences at Duke and the senior author of the study. "Many biology texts today still refer to the appendix as a vestigial organ." Furthermore, over 70% of all primate and rodent taxonomic families contain species with an appendix.14 The vestigial organ idea helped fool millions of people into believing the theory of evolution. Today, this "proof" is down the toilet.
Violating the lawThe theory of Evolution violates two laws of science. The Second Law of Thermodynamics (law of increasing entropy) says that things which start out concentrated together spread out over time. If you heat one room in a house, then open the door to that room, eventually the temperature in the whole house evens out (reaches equilibrium). Knowing how far this evening-out has progressed at any point in time tells you the entropy. Entropy can measure the loss of a system's ability to do work. Entropy is also a measure of disorder, and that is where evolution theory hits an impenetrable wall. Natural processes proceed in only one direction, toward equilibrium and disorder. Things fall apart over time, they do not get more organized. We can overcome this by making a machine and adding energy, but the Second Law prevents such a machine from assembling spontaneously from raw materials.
The Law of Biogenesis was established by Louis Pasteur three years after Darwin's book was published, and simply says that life only comes from life. Living cells divide to make new cells, and fertilized eggs and seeds develop into animals and plants, but chemicals never fall together and life appears. Evolutionists often call certain chemicals "the building blocks of life", giving people the false impression that you just stack the building blocks together and you get life. No one has ever done that, including the famous 1953 Miller/Urey experiment where all they got were clumps of amino acids. Many people mistakenly think scientists have made life from chemicals in the lab, but they have not (though many have tried very hard). If one were to succeed, you would know about it. He would get every science award there is, be all over the news, and have movies, books, buildings, statues, and schools dedicated to him, so desperate are evolutionists on this matter. For something to be a law of science, it can never be found to have been violated, even once, over thousands of trials. No exceptions. A theory that violates two laws of science is in big trouble.
When confronted with the Second Law of Thermodynamics, evolutionists usually use two tricks to try to escape. The first is to state that "it only applies to closed systems, and biological creatures are open systems, so it doesn't affect evolution" (they actually intend to say isolated, not closed, but we know what they mean). The fact is that the Second Law applies to all systems, open or closed, and to all actions and chemical reactions, from molecules to galaxies. The words "except for..." are not in this universal law. A thermodynamics system is simply any part of the universe we want to study. If we are doing an experiment in a bottle, the inside of the bottle is our system and the bottle itself is the "walls" of the system. There are only 3 kinds of systems: if no energy or matter can pass through the walls, it is an isolated system; if energy can pass through but matter cannot, it is a closed system; if both energy and matter can pass through the walls, it is an open system. Now, it is true that the laws of thermodynamics and entropy are defined in terms of isolated systems, because that is the simplest way to express them. However, experts who write textbooks on the subject are quick to say that isolated systems do not occur in nature. For practical applications, a procedure called the Legendre Transform mathematically converts entropy to a variable called Gibbs free energy that is useful for working with real-world systems. Most natural systems are open, but it is convenient to model them as closed. For example, even though a bacterium is an open system, modeling it as a closed system makes it easier to understand chemical reactions in it.2,8
You are an open system. You eat food (which comes from outside yourself) and your body survives and grows. Evolutionists believe that all we need is an open system with sufficient energy flowing into it for evolution to succeed. If that were so, you could just stand right behind a jet engine as the aircraft prepares for takeoff, absorb that blast of energy, and evolve to a higher life form. In reality, of course, you would be incinerated because absorbing energy without a mechanism to convert it to a useful form and employ it is destructive or useless. The mechanism must be very specific. Sticking food in your ear will not work; it must go into your mouth and through the digestive system. And the mechanism must be in place and functioning first, before energy is added, or the energy is wasted. The "closed system" ploy is just an attempt to avoid dealing with the Second Law because the Law prohibits any functioning biological mechanism from falling together by pure chance, without assistance or plan, using only the properties of matter.
The second trick they use is to say that "when you freeze water, the disordered molecules become beautifully ordered ice crystals or snowflakes. If water can bypass the Second Law and organize its molecules by a natural process, why not the chemicals of life?" At room temperature, water molecules are bouncing off each other and you have water. When you take away heat and they freeze, water molecules stick to each other with weak molecular bonds, forming ice crystals and snowflakes because of the shape of the H2O molecule. The same thing happens if you put a bunch of weak magnets in a jar and shake it. The magnets bounce around. When you stop, the magnets stick together. They are at a lower energy level. There is order, yet no complexity - just a simple repetitive structure that does not do anything. The Second Law is not bypassed or violated.
But guess what. Amino acid molecules that form proteins, and nucleotide molecules that form DNA and RNA resist combining at any temperature. To combine, they need the help of mechanisms in a living cell or a biochemist in an organic chemistry laboratory.18 It means that nothing happens in the primeval soup, the pond of chemicals where evolutionists believe life began.
DNA is made of only right-handed versions of nucleotides, while proteins are made of only left-handed versions of amino acids. Yet any random chemical reaction that produced nucleotides or amino acids would make an equal mix of left and right-handed versions of each. Even if the thousands of nucleotides needed to form a DNA molecule, or the hundreds of amino acids needed to form a protein molecule were able to combine from the mix, they would be a jumble of left and right-handed versions that could not function at all. This is the problem of "chirality", and evolutionists have never been able to solve it.
Ilya Prigogene coauthored a paper in 1972 that says in an open "system there exists a possibility for formation of ordered, low-entropy structures at sufficiently low temperatures. This ordering principle is responsible for the appearance of ordered structures such as crystals... Unfortunately this principle cannot explain the formation of biological structures."36 Prigogene won the Nobel Prize in Chemistry in 1977 for research on dissipative structures, such as tornados, for contributions to nonequilibrium thermodynamics, and for bridging the gap between biology and other sciences. Evolutionists wrongly claim he won for showing how thermodynamics could explain the formation of organized systems, from fluctuations in chaos, that lead to the origin of life. They thought he was their hero. Over thirty-five years later, nothing has come of it. There is no escape from the Second Law of Thermodynamics. It prohibits the spontaneous origin of life and macroevolution.
Mindboggling complexity - proteinsEven a single cell is not simple. In Darwin's day researchers looked at cells under the microscope and saw little balloons filled with goo they called protoplasm, so they thought cells were simple forms of life. 150 years later we know that there are many types of cells, and each cell is a little city at work. The smallest known genome (Mycoplasma genitalium) has 482 genes.19 The minimum possible for an organism to survive is probably 200 to 300 genes. Most bacteria have 1000 to 4000 genes. A popular textbook on the cell1 is 1600 pages long and weighs 7 pounds. Everything about the cell is stunningly complex. Plants and animals contain a great variety of cells. The human body has about 210 different types of cells.
Cells are made of proteins, and everything that goes on in a creature involves proteins interacting with each other. Proteins are generally 50 to 2000 amino acids long; a typical one has about 300 amino acids.1 Ribosomes are molecular machines that build proteins in cells, using messenger RNA as the template. Here is an overview of how a bacterial ribosome "translates" RNA into protein. Every protein in bacteria is made this way.
From: Schmeing, T. Martin, V. Ramakrishnan. 29 October 2009. What recent ribosome structures have revealed about the mechanism of translation. Nature, Vol. 461, pp. 1234-1242.

|
Diagram of a folded protein
|
Origami
|
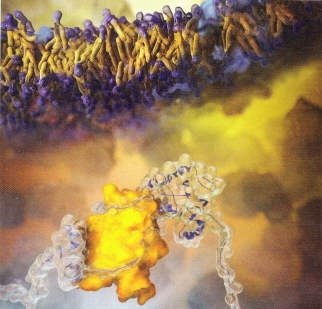
"A chaperone protein (bottom, yellow) called SecB guides the folding of another protein (transparent) in this artist's illustration."--Science News, December 1, 2007, Vol. 172, p. 342
- Replication, recombination, and repair
- Transcription
- Cell cycle control, mitosis, and meiosis
- Defense mechanisms
- Cell wall/membrane biogenesis
- Signal transduction mechanisms
- Intracellular trafficking and secretion
- Translation
- Post-translational modification, protein turnover, chaperones
- Energy production and conversion
- Carbohydrate transport and metabolism
- Amino acid transport and metabolism
- Nucleotide transport and metabolism
- Coenzyme transport and metabolism
- Lipid transport and metabolism
- Inorganic ion transport and metabolism
- Secondary metabolite biosynthesis, transport, and catabolism
So evolutionists have to believe that for each protein, pure chance laid out long strings of amino acids that fold themselves into the exact shapes needed to interact with other specialized proteins and, where needed, get help from chaperone proteins which themselves appeared by chance. The necessary proteins cannot be invented one at a time. Either they are all there, ready to work together, or nothing happens and they disintegrate. Yet even if it could design proteins, mutation-natural selection would only work on one at a time sporadically over many years. Considering just the complexity of proteins, the notion of creating them with mutation-natural selection is as silly as asking someone to build a television set with a spoon and a toothbrush. If Darwin had known what we have learned about proteins, he probably would have abandoned the theory of evolution.
Darwin
himself wrote in chapter 6 of On the Origin of Species
that "natural selection can act only by the
preservation and accumulation of infinitesimally small inherited
modifications, each profitable to the preserved being... If it could
be demonstrated that any complex organ existed, which could not
possibly have been formed by numerous, successive, slight modifications,
my theory would absolutely break down."
Duplicate
genesDo evolutionists admit defeat? Never! They temporarily sidestep natural selection, saying the mutations in DNA needed to build a complicated new part quietly accumulate in duplicate genes, because by themselves each of the necessary mutations is neutral, neither beneficial nor harmful. Then, millions of years later, all are in place. The new part starts working, natural selection chooses it, and the improved creature is off to the races. This scenario exists only in the mind of the evolutionist.
It is true that just one copy of a gene needs to work right. There are many duplicate genes in DNA, and the idea is that they are free to change without hurting the creature.
But the duplicates are also protected from evolution theory's only "guiding force", natural selection. Any novel genes would have to mesh with the elaborate regulatory networks of living creatures, and do so completely randomly, without harmful mutations.
A review of many experiments with duplicate genes found that "duplication can and does facilitate important adaptations by tinkering with existing compounds". The "random mutations and recombinations considered were observed to tweak, tinker, copy, cut, divide, and shuffle existing genetic information around, but fell short of generating genuinely distinct and entirely novel functionality." "Gene duplication results in the copying and preservation of biological information, and not its transformation as something original." It is "insufficient in explaining the origination of the highly complex information pertinent to the essential functioning of living organisms." --Bozorgmehr, Joseph Esfandiar Hannon. 22 December 2010. Is Gene Duplication a Viable Explanation for the Origination of Biological Information and Complexity? Complexity, published online, DOI 10.1002/cplx.20365, pp. 1-14.
Repairing mutations
So duplicate genes are not macroevolution's secret laboratory. Furthermore, everyone agrees that harmful mutations appear many, many times more often than mutations needed for new construction ever could. Over those millions of years, slightly harmful mutations that are hidden, or not destructive enough for natural selection to remove, would also quietly accumulate. This would produce creatures loaded up with highly polluted genes. Survival of the barely functional? We do not find this either because cells have mechanisms that maintain the original design of a creature within its variation boundaries, and minimize the accumulation of mutations. These include:
- A proofreading system that catches almost all errors
- A mismatch repair system to back up the proofreading system
- Photoreactivation (light repair)
- Removal of methyl or ethyl groups by O6 - methylguanine methyltransferase
- Base excision repair
- Nucleotide excision repair
- Double-strand DNA break repair
- Recombination repair
- Error-prone bypass40
Genome size
Scientists have found that the number of genes a creature has is not a good measure of how complex it is. For example, the human genome is 23 times larger than the fruit fly genome (3.2 billion base pairs versus 137 million), yet humans have less than twice the number of protein coding genes (21,000 versus 13,000) . Yeast has about 6,000 genes.
 |
|
The tiny water flea
Daphnia pulex has more genes than humans do;
up to 39,000 at last count. Water
Flea Boasts Whopper Gene Count. 5 June 2009. Science,
Vol. 324, No. 5932, p. 1252. |
|
|
|
|
 |
|
So does the pea aphid
Acyrthosiphon pisum,
with 34,600.Water
Flea Boasts Whopper Gene Count. 5 June 2009. Science,
Vol. 324, No. 5932, p. 1252. |
Junk DNA
Only a small portion of a creature's DNA is protein-coding genes (around 3% in humans). In the 1970s, evolutionists began calling the rest of it "junk DNA", saying this collection of useless evolutionary debris showed there was no intelligent design involved. Decades later, researchers are finding that the "junk" does vital work. Some of this DNA plays a role in turning genes on and off at the right moments in a developing embryo29. Other bits separate coding and regulating sections, like punctuation marks in writing, so that DNA is not a long run-on sentence30. Other bits called Alu elements, found only in primates, can be spliced in or out during RNA processing to make different versions of the same gene.31 The "junk" label discouraged research into this part of the genome for many years; who would want to waste their time studying it? An article in the 7 September 2012 issue of Science was titled "ENCODE project writes eulogy for junk DNA". "This week, 30 research papers... sound the death knell for the idea that our DNA is mostly littered with useless bases. A decadelong project, the Encyclopedia of DNA Elements (ENCODE), has found that 80% of the human genome serves some purpose". "The ENCODE effort has revealed that a gene's regulation is far more complex than previously thought, being influenced by multiple stretches of regulatory DNA located both near and far from the gene itself and by strands of RNA not translated into proteins, so-called noncoding RNA."-- Pennisi, Elizabeth. 7 September 2012. Science, Vol. 337, pp. 1159-1161. The ENCODE project is not the last word on the genome. As time goes on, expect researchers to find that more and more of the remaining genome is vital information code for the construction and function of humans.
Networks
The living things of the world are extremely varied and intricately made, yet the theory of evolution has always been about simplicity: once upon a time, some chemicals assembled, began to make copies of themselves, and little by little changed into all life forms. Evolutionists like to use the words "simply" and "merely" when telling their stories to the public. There is certainly nothing complicated about the idea of mutation-natural selection. However, by the year 2000, research had reached a point where a new branch of biology was needed: Systems Biology. Discoveries in this field are the exact opposite of merely simple. Biological systems are vastly more complex than anyone could imagine. Some wonder if we will ever fully understand them.

A small section of a biological system in an organism, displayed as a 3D network
"The interesting point coming out of all these studies is how complex these systems are; the different feedback loops and how they cross-regulate each other and adapt to perturbations are only just becoming apparent. The simple pathway models are a gross oversimplification of what is actually happening", says Mike Tyers, a systems biologist at the University of Edinburgh, UK.-- Blow, Nathan. 16 July 2009. Untangling the protein web. Nature, Vol. 460, pp. 415-418.
This is a map of how the genes in a cell of the budding yeast Saccharomyces cerevisiae interact with one another. Each color shows what a group of genes does. Genes in these functional networks interact with other genes throughout the cell; a cell of yeast.
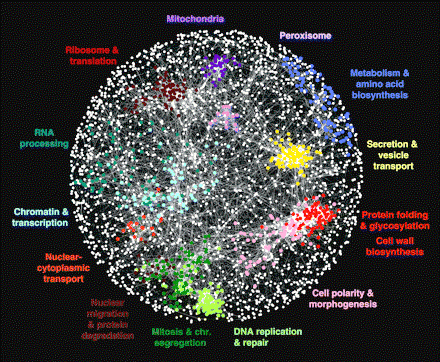
Costanzo, Michael, et al. 22 January 2010. The Genetic Landscape of a Cell.
Science, Vol. 327, No. 5964, pp. 425-431.
An embryo has a particular growth program for the type of creature it will develop into. Yet it is likely that all the different programs are constructed from just a few types of sub-circuits. "Structurally similar sub-circuits, but composed of different regulatory genes" do "similar developmental jobs in different GRNs." This discovery gives researchers hope that they can use these "modules of developmental logic function" to decipher the "enormous mazes of interconnections in system level GRNs". You can tell what a sub-circuit does by its shape or structure. There is a sub-circuit for each task, and GRNs are made up of sub-circuits. The same control processes are used "throughout embryonic development because the problems that have to be solved are general: the initial spatial inputs have to be interpreted, the regulatory state then has to be locked down (the initial inputs are always transient), signals have then to be generated, other states have to be excluded, and differentiation drivers have to be activated. It is not surprising that all this requires a lot of sequential circuitry."
GRNs in embryos "are hierarchical in their overall structure. Their depth reflects the long sequence of regulatory steps required to complete any component of embryonic development." A GRN might have many layers of sub-circuits or very few, depending on its job. The last step in the chain of command is the signal for "batteries" (groups) of genes to change stem cells into specific types of cells (such as muscle, blood, nerve, etc.) at the right place.-- Davidson, Eric H. 16 December 2010. Emerging properties of animal gene regulatory networks. Nature, Vol. 468, pp. 911-920.
Some evolutionists have publicly welcomed GRNs because a change in one controller can affect many genes, and we are back to simplicity. That is like saying a child can use Windows 7 operating system on a computer. Just point and click with a mouse, and the computer does many complicated things. It is simplicity itself. So why are GRNs the death blow to evolution theory? It took computer and software engineers decades of intelligent design to build the computer and Windows 7. GRNs are the operating system of living things. The theory of evolution cannot explain how gene regulatory networks came to be. As with other insurmountable problems with the theory, this one remains in their pile marked "needs further study".
Today there is an explosion of knowledge going on in the study of gene regulatory networks. But it is not led, assisted, or even inspired by the theory of evolution. "We have little empirical knowledge on the evolutionary history of such networks."-- Dean, Antony M., Joseph W. Thornton. September 2007. Mechanistic approaches to the study of evolution: the functional synthesis. Nature Reviews Genetics, Vol. 8, pp. 675-688.
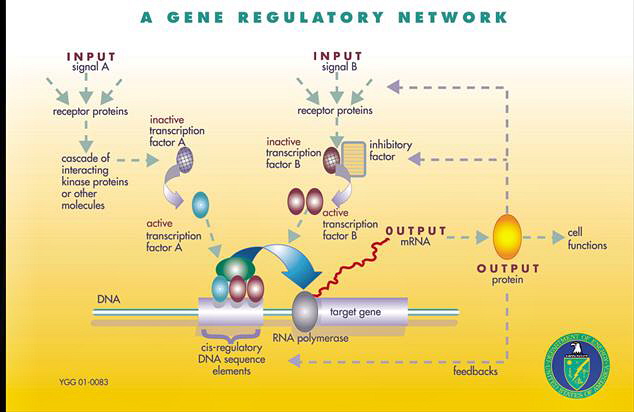
- Specialized GRNs determine which genes are active or inactive in each part of a developing creature
- GRN sub-circuits, usually consisting of 3 to 8 regulatory genes plus the elements they regulate, perform specific functions
- Switches permit or forbid the activity of whole sub-circuits
- Gene batteries are groups of genes required for particular cell functions; they are controlled by a small set of transcriptional drivers
- Segments of DNA a few hundred base pairs long, called cis-regulatory elements, control expression of the genes near them
- Signals are deployed between one cell and another using cis-regulatory elements
Mutation-natural selection could no more build the vast, intricate networks in living creatures than a beaver could build the Hoover dam.

To the next levelAt this point we are light-years beyond the simplistic
notions of Darwinism. Now even systems
biology is being overwhelmed.
"The protein p53, for example, was discovered in
1979." "It soon gained notoriety as a
tumor suppressor - a 'guardian of the genome' that stifles cancer growth by
condemning genetically damaged cells to death. Few proteins have been studied more than p53."
"Researchers now know that p53 binds to thousands of sites
in DNA, and some of these sites are thousands of base pairs away from any
genes. It influences cell growth,
death, structure and DNA repair. It
also binds to numerous other proteins." "Through a process known as alternative splicing, p53 can take nine
different forms, each of which has its own activities and chemical
modifiers. Biologists are now realizing
that p53 is also involved in processes beyond cancer, such as fertility and
very early embryonic development."
Research "has dismantled old ideas about signaling 'pathways', in which proteins such as p53 would trigger a defined set of
downstream consequences. 'When we started
out, the idea was that signaling pathways were fairly simple and linear,' says
Tony Pawson, a cell biologist at the University of Toronto in Ontario. 'Now, we appreciate that the signaling
information in cells is organized through networks of information rather than
simple discrete pathways. It's
infinitely more complex.'
"
"Systems biology was supposed to help scientists make sense
of the complexity. The hope was that by
cataloguing all the interactions in the p53 network, or in a cell, or between a
group of cells, then plugging them into a computational model, biologists would
glean insights about how biological systems behaved."
Unfortunately, "there is no way to gather all the relevant
data about each interaction". "In many
cases, the models themselves quickly become so complex that they are unlikely
to reveal insights about the system, degenerating instead into mazes of
interactions". "Many of the mechanisms
and principles governing inter- and intracellular behavior are still a
mystery."-- Hayden, Erika Check. 1 April
2010. Life is Complicated. Nature, Vol. 464, pp. 664-667.
When
cells repair damaged DNA using so-called "replicate DNA"
(for making copies) or "transcribe DNA" (the first step
in making a protein), the parts are rapidly assembled from a pool
of parts floating in the nucleus of a cell to form "factories".
The size of a repair center is according to the amount of
damage it has to repair. "A replication factory persists
for a few minutes before it disassembles. A new factory is
then assembled... immediately adjacent to the previous one".
Whether it is repair, replication, or transcription, once
the job is done the "factories" disassemble and the parts
float back into the pool.32
Chromatin is DNA packaged into
chromosomes. Chromatin is in constant motion.3
Different sections along DNA are apparently guided to each
other directly and rapidly, forming loops.11
Chromatin loops are very common. Loops range in size
from thousands to hundreds-of-thousands of bases long. Loops
bring together genes and their regulators to form "transcription
hubs" where transcription can occur.32
"Long-range interactions can occur over very large genomic
distances, up to tens of megabases". "Interactions
occur not only along chromosomes, but also between them." "Chromosomes
extensively interact with each other"42,
and neighboring chromosomes intermingle.32There is a "bewildering complexity in long-range communication among a variety of genomic elements across chromosomes and the genome."42
The Bottom Line
Evolutionists assume evolution is true, then write endlessly about when and where it happened, rates and lineages, etc. But if macroevolution is physically impossible in the real world, and it is, then all the rest is fantasy. There are only two possibilities. Either every part of every living thing arose by random chance, or an intelligence designed them.
 |
|
It is now clear that the theory of evolution's only mechanism for building
new parts and creatures, mutation-natural selection, is totally,
utterly, pathetically inadequate. |
Many scientists are with us
The only tactic left to evolutionists is to ridicule their critics as simpletons who don't understand how their pet theory really works. Here is a link to a roster of hundreds of professionals whose advanced academic degrees certify that they thoroughly understand evolution theory. They also have the courage to defy the high priests of academia by voluntarily adding their names to a skeptics list against Darwinism.
Some revealing quotes
Philip S. Skell, a member of the National Academy of Sciences, wrote in the August 29, 2005 edition of The Scientist: "I recently asked more than seventy eminent researchers if they would have done their work differently if they had thought Darwin's theory was wrong. The responses were all the same: No. I also examined the outstanding discoveries of the past century: the discovery of the double helix; the characterization of the ribosome; the mapping of genomes; research on medications and drug reactions; improvements in food production and sanitation; the development of new surgeries; and others. I even queried biologists working in areas where one would expect the Darwinian paradigm to have most benefited research, such as the emergence of resistance to antibiotics and pesticides. Here, as elsewhere, I found that Darwin's theory had provided no discernible guidance, but was brought in, after the breakthroughs, as an interesting narrative gloss."--Philip S. Skell. August 29, 2005. Why Do We Invoke Darwin? The Scientist, Vol. 19, No. 16, p. 10.
- Philip S. Skell was Evan Pugh Professor of Chemistry, Emeritus at Penn State University. He is sometimes called "the father of carbene chemistry" in organic chemistry, and is widely known for the "Skell Rule", which was first applied to carbenes - the "fleeting species" of carbon. The rule, which predicts the most probable pathway through which certain chemical compounds will be formed, found use throughout the pharmaceutical and chemical industries. He said that during World War II "I was personally associated with an antibiotics research group, engaged in the full range of activities, from finding organisms which inhibited bacterial growth to the isolation and proof of structure of the antibiotics they produced." Professor Skell died Nov. 21, 2010.
A. Clark, R.W. 1985. The Life of Ernst Chain: Penicillin and Beyond. New York: St. Martin's Press, p. 147.
B. Chain, E. 1970. Social Responsibility and the Scientist in Modern Western Society (Robert Waley Cohen memorial lecture). London: The Council of Christians and Jews, p. 25.
"My experiences with science led me to God. To be forced to believe only one
conclusion -- that everything in the universe happened by chance -- would violate the
very objectivity of science itself. Certainly
there are those who argue that the universe evolved out of a random process,
but what random process could produce the brain of a man or the system of the
human eye? Some people say that science
has been unable to prove the existence of a Designer... They challenge science to prove the existence
of God. But, must we really light a candle to see the sun?" --Wernher
von Braun 1912 - 1977
- "Wernher von Braun is, without doubt, the greatest rocket scientist in history. His crowning achievement, as head of NASA's Marshall Space Flight Center, was to lead the development of the Saturn V booster rocket that helped land the first men on the Moon in July 1969." --From NASA's webpage: http://earthobservatory.nasa.gov/Features/vonBraun/
Sean B. Carroll, of the Medical Institute and Laboratory of Molecular Biology at the University of Wisconsin--Madison, wrote in a 2001 edition of Nature: "A long-standing issue in evolutionary biology is whether the processes observable in extant populations and species (microevolution) are sufficient to account for the larger-scale changes evident over longer periods of life's history (macroevolution). Outsiders to this rich literature may be surprised that there is no consensus on this issue."-- Sean B. Carroll. 8 February 2001. Nature, Vol. 409, p. 669.
A symposium on evolution was held at the European Molecular Biology Laboratory in Heidelberg, Germany in November 2001, organized by PhD students. The meeting report says that "the symposium ended with a panel discussion about questions of microevolution (evolution within the species) and macroevolution (evolution after speciation). The issue at stake was whether extrapolation from the selection theory operating on organisms is sufficient to explain all patterns of macroevolution. In other words, do we need an independent body of theory to explain the changes occurring above, as opposed to at, the species level? There was no general agreement among the panel members. It seems that the jury is still out on this important question."-- Gaspar Jekely. 2002. Meeting report - Evolution in a nutshell. European Molecular Biology Organization reports, Vol. 3, No. 4, pp. 307-311.
"Biology has been re-integrated twice already, first by Darwin in 1859 and then during the 'Modern Synthesis' of the 1920s and 1930s. In both cases, the success of these syntheses rested in part on ignorance. Charles Darwin could reasonably integrate biology in the 19th Century on a relatively elegant evolutionary foundation partly because a great deal was not yet known about cellular and biochemical machinery." "Like Darwin's synthesis, the form of the Modern Synthesis was shaped in part by ignorance of important features of life that were at the time unknown to science. Specifically, the molecular biology of the cell remained largely unknown." "The view of life that most biologists had from 1935 to 1965 was highly simplified. Some of the assumptions at the foundation of the Modern Synthesis started to crumble in the 1970s. Common mid-20th Century assumptions about how cells, organisms, and species work have thus been undermined." "This might seem like reason for despair about the future of biology, but there are two mitigations to consider. First, this complexity was always there. Darwin and many later biologists realized that their simple models were erected like piers over swampy ground. They just didn't know how deep the muck was. Second, we now have powerful genomic tools for addressing complex phenomena throughout biology." "Some may feel that the view of life supplied by nascent 21st Century biology is painfully complicated, if not perverse. For our part, we think that the historical complexity and versatility that we now know to characterize life are inspiring and challenging."
"Origin of Life" research, continued
The theory of evolution says life started from raw chemicals. Evolutionists long ago handed the problem off to specialists, trusting that they would come up with something. The specialists have spent many frustrating decades trying to figure out how DNA assembled itself. They have two approaches to the problem, and those on one side think the other side is wrong. Here is the essence of both views, synthesized from two research papers:
"The conceivable paths toward life's emergence have been dominated by two fundamentally different views in origin-of-life research: the genetics- or replication-first approach, and the metabolism-first scenario. Both schools acknowledge that the critical requirement for primitive evolvable systems (in the Darwinian sense) is to solve the problems of information storage and reliable information transmission. Disagreement starts, however, in the way information was first stored. All present life is based on digitally encoded information."V The mainstream prebiotic evolutionary scenario is the "RNA world".S "Textbooks often assert that life began with specialized complex molecules, such as RNA, that are capable of making their own copies. This scenario has serious difficulties, but an alternative has remained elusive."S "We do not know how the transition to digitally encoded information has happened in the originally inanimate world; that is, we do not know where the RNA world might have come from."V
"An alternative appears to be necessary for the RNA-centric paradigm of the origin of life."S "No known cellular constituent is capable of self-replication in pure form. Even DNA is absolutely dependent on other cellular components for making its own copies."S "One is compelled to consider an alternative: that self-replication has never been a property of individual molecules, but rather one of molecular ensembles."S "The crucial origin of life question then becomes how natural selection was initiated by some molecular assortments, irrespective of their exact chemistry."S "Life on our planet could have begun as a random chemistry melting pot, a 'garbage-bag world' with myriads of different chemical configurations."S "A complex chain of evolutionary events, yet to be deciphered, could then have led to the common ancestors of today's free-living cells, and to the appearance of DNA, RNA and protein enzymes."S
"Was a network of chemical reactions able to increase in complexity and eventually undergo Darwinian selection?"V "We demonstrate here that replication of compositional information is so inaccurate that fitter compositional genomes cannot be maintained by selection and, therefore, the system lacks evolvability."V "The computed population dynamics of growing noncovalent molecular assemblies that undergo splitting when a critical size is reached clearly illustrates that compositional assemblies do not evolve."V "We conclude that this fundamental limitation of ensemble replicators cautions against metabolism-first theories of the origin of life."V "We now feel compelled to abandon compositional inheritance as a jumping board toward real units of evolution."V
S -- Segre, Daniel, Doron Lancet. 2000. Composing life. European Molecular Biology Organization (EMBO) Reports, Vol. 1, No. 3, pp. 217-222.
V -- Vasas, Vera, Eors Szathmary, Mauro Santos. January 26, 2010. Lack of evolvability in self-sustaining autocatalytic networks constraints metabolism-first scenarios for the origin of life. Proceedings of the National Academy of Sciences of the United States of America (PNAS), Vol. 107, No. 4, pp. 1470-1475.
So neither approach works.
Here are excerpts from candid reports by two scientists who have spent many years in "origin of life" research. These men support evolution, but insist that experimental evidence back up every claim.
This is "what has been called the NASA definition of life: Life is a self-sustained chemical system capable of undergoing Darwinian evolution." "Richard Dawkins elaborated on this image of the earliest living entity in his book The Selfish Gene: 'At some point a particularly remarkable molecule was formed by accident. We will call it the Replicator. It may not have been the biggest or the most complex molecule around, but it had the extraordinary property of being able to create copies of itself.' When Dawkins wrote these words 30 years ago, DNA was the most likely candidate for this role." "Unfortunately... DNA replication cannot proceed without the assistance of a number of proteins". So "which came first, the chicken or the egg? DNA holds the recipe for protein construction. Yet that information cannot be retrieved or copied without the assistance of proteins. Which large molecule, then, appeared first in getting life started--proteins (the chicken) or DNA (the egg).?"
"A possible solution appeared when attention shifted to a new champion--RNA." According to this view, "life began with the appearance of the first RNA molecule. In a... 1986 article, Nobel Laureate Walter Gilbert of Harvard University wrote in the journal Nature: 'One can contemplate an RNA world, containing only RNA molecules that serve to catalyze the synthesis of themselves. The first step of evolution proceeds then by RNA molecules performing the catalytic activities necessary to assemble themselves from a nucleotide soup.' In this vision, the first self-replicating RNA that emerged from non-living matter carried out the functions now executed by RNA, DNA and proteins." "Perhaps two-thirds of scientists publishing in the origin-of-life field... still support the idea that life began with the spontaneous formation of RNA or a related self-copying molecule."
"How did that first self-replicating RNA arise?" Most people know of an "experiment published in 1953 by Stanley Miller. He applied a spark discharge to a mixture of simple gases that were then thought to represent the atmosphere of the early Earth. Two amino acids of the set of 20 used to construct proteins were formed in significant quantities, with others from that set present in small amounts." "Some writers have presumed that all of life's building blocks could be formed with ease in Miller-type experiments and were present in meteorites and other extraterrestrial bodies. This is not the case."
"A careful examination of the results of the analysis of several meteorites led the scientists who conducted the work to a different conclusion: inanimate nature has a bias toward the formation of molecules made of fewer rather than greater numbers of carbon atoms, and thus show no partiality in favor of creating the building blocks of our kind of life." "RNA's building blocks, nucleotides, are complex substances as organic molecules go." "Amino acids, such as those produced or found in these experiments, are far less complex than nucleotides". "No nucleotides of any kind have been reported as products of spark discharge experiments or in studies of meteorites."
"To rescue the RNA-first concept from this otherwise lethal defect, its advocates have created a discipline called prebiotic synthesis. They have attempted to show that RNA and its components can be prepared in their laboratories in a sequence of carefully controlled reactions." Finding "a specific organic chemical in any quantity... would justify its classification as 'prebiotic,' a substance that supposedly had been proved to be present on the early Earth. Once awarded this distinction, the chemical could then be used in pure form, in any quantity, in another prebiotic reaction. The products of such a reaction would also be considered 'prebiotic' and employed in the next step in the sequence." "Unfortunately, neither chemists nor laboratories were present on the early Earth to produce RNA." "The analogy that comes to mind is that of a golfer, who having played a golf ball through an 18-hole course, then assumed that the ball could also play itself around the course in his absence. He had demonstrated the possibility of the event; it was only necessary to presume that some combination of natural forces (earthquakes, winds, tornadoes and floods, for example) could produce the same result, given enough time."
"Many chemists, confronted with these difficulties, have fled the RNA-first hypothesis as if it were a building on fire. One group, however, still captured by the vision of the self-copying molecule, has opted for an exit that leads to similar hazards. In these revised theories, a simpler replicator arose first and governed life in a 'pre-RNA world.' Variations have been proposed in which the bases, the sugar or the entire backbone of RNA have been replaced by simpler substances, more accessible to prebiotic syntheses. Presumably, this first replicator would also have the catalytic capabilities of RNA. Because no trace of this hypothetical primal replicator and catalyst has been recognized so far in modern biology, RNA must have completely taken over all of its functions at some point following its emergence."
"Further, the spontaneous appearance of any such replicator without the assistance of a chemist faces implausibilities that dwarf those involved in the preparation of a mere nucleotide soup. Let us presume that a soup enriched in the building blocks of all of these proposed replicators has somehow been assembled, under conditions that favor their connection into chains. They would be accompanied by hordes of defective building blocks, the inclusion of which would ruin the ability of the chain to act as a replicator." "There is no reason to presume that an indifferent nature would not combine units at random".
"Probability calculations could be made, but I prefer a variation on a much-used analogy. Picture a gorilla (very long arms are needed) at an immense keyboard connected to a word processor. The keyboard contains not only the symbols used in English and European languages but also a huge excess drawn from every other known language and all of the symbol sets stored in a typical computer. The chances for the spontaneous assembly of a replicator in the pool I described above can be compared to those of the gorilla composing, in English, a coherent recipe for the preparation of chili con carne. With similar considerations in mind, Gerald F. Joyce of the Scripps Research Institute and Leslie Orgel of the Salk Institute concluded that the spontaneous appearance of RNA chains on the lifeless Earth 'would have been a near miracle.' I would extend this conclusion to all of the proposed RNA substitutes that I mentioned above." "Nobel Laureate Christian de Duve has called for 'a rejection of improbabilities so incommensurably high that they can only be called miracles, phenomena that fall outside the scope of scientific inquiry.' DNA, RNA, proteins and other elaborate large molecules must then be set aside as participants in the origin of life."
What is left? Theories that "employ a thermodynamic rather than a genetic definition of life, under a scheme put forth by Carl Sagan in the Encyclopedia Britannica: A localized region which increases in order (decreases in entropy) through cycles driven by an energy flow would be considered alive." "I estimate that about a third of the chemists involved in the study of the origin of life subscribe to theories based on this idea."
It requires: "1) A boundary... to separate life from non-life." "2) An energy source". "3) A coupling mechanism must link the release of energy to the organization process that produces and sustains life. The release of energy does not necessarily produce a useful result. Chemical energy is released when gasoline is burned within the cylinders of my automobile, but the vehicle will not move unless that energy is used to turn the wheels. A mechanical connection, or coupling, is required." "4) A chemical network must be formed, to permit adaptation and evolution" "on a path that leads to increased organization." "5) The network must grow and reproduce." "We can imagine, on the early Earth, a situation where many startups of this type occur, involving many alternative driver reactions and external energy sources. Finally, a particularly hardy one would take root and sustain itself." "A system of reproduction must eventually develop." "Once independent units were established, they could evolve in different ways and compete with one another for raw materials; we would have made the transition from life that emerges from nonliving matter through the action of an available energy source to life that adapts to its environment by Darwinian evolution." "Many further steps in evolution would be needed to 'invent' the elaborate mechanisms for replication and specific protein synthesis that we observe in life today." They "would not reveal the specific events that led to the familiar DNA-RNA-protein-based organisms of today."
"Systems of the type I have described usually have been classified under the heading 'metabolism first', which implies that they do not contain a mechanism for heredity. In other words, they contain no obvious molecule or structure that allows the information stored in them (their heredity) to be duplicated and passed on to their descendants." "Over the years, many theoretical papers have advanced particular metabolism first schemes, but relatively little experimental work has been presented in support of them." "They have not yet demonstrated the operation of a complete cycle or its ability to sustain itself and undergo further evolution. A 'smoking gun' experiment displaying those three features is needed to establish the validity of the small molecule approach."
Shapiro, Robert. June 2007. A Simpler Origin for Life. Scientific American, Vol. 296, pp. 24-31.
Robert Shapiro, Ph.D. Harvard, is professor emeritus of chemistry and senior research scientist at New York University. He is author or co-author of over 125 publications, primarily in the area of DNA chemistry. In 2004 he was awarded the Trotter Prize in Information, Complexity and Inference. Shapiro has been involved in the search for origin of life mechanisms, and has written four books on the subject for the general public.
"The feasibility of any particular proposed prebiotic cycle must depend on arguments about chemical plausibility, rather than on a decision about logical possibility." The metabolic cycles that have been identified by biochemists are of two kinds: simple cycles and autocatalytic cycles. The citric acid cycle" is an example of a simple cycle. "The reverse citric acid cycle" is an example of an autocatalytic cycle. "Each molecule of citric acid introduced into the cycle results... in the generation of two molecules of citric acid." "That is why the cycle is described as autocatalytic." "The proposal that the reverse citric acid cycle operated... on the primitive Earth has been a prominent feature of some scenarios for the origin of life."
"A different kind of autocatalytic cycle, which has no analog in biochemistry, has been hypothesized by Stuart Kauffman to self-organize spontaneously whenever amino acids condense together to form peptides." "Could prebiotic molecules and catalysts plausibly have the attributes... to make the self-organization of the cycles possible?"
"The identification of a cycle of plausible prebiotic reactions is a necessary but not a sufficient step toward the formulation of a plausible self-organizing prebiotic cycle. The next, and more difficult step, is justifying the exclusion of side reactions that would disrupt the cycle." "It is not completely impossible that sufficiently specific mineral catalysts exist for each of the reactions of the reverse citric acid cycle, but the chance of a full set of such catalysts occurring at a single locality on the primitive Earth in the absence of catalysts for disruptive side reactions seems remote in the extreme."
"It has sometimes been implied or claimed that [autocatalytic] cycles are not only stable, but also are capable of evolving to form nonenzymatic networks of great complexity. Genetic materials are then seen as late additions to already fairly complex evolved life forms. According to this view, a genetic material merely adds stability to systems that already have a substantial 'information content'. "
"One way of achieving something useful might be to use one of the constituents of the core cycle as the starting point of a second independent autocatalytic cycle." "Another suggestion that might be explored is the possibility of a side reaction generating a catalyst for one of the reactions of the core cycle." "However, neither of these possibilities, nor any others with which I am familiar explains how a complex, interconnected family of cycles capable of evolution could arise or why it should be stable." "What is essential, therefore, is a reasonably detailed description, hopefully supported by experimental evidence, of how an evolvable family of cycles might operate." "Without such a description, acceptance of the possibility of complex nonenzymatic cyclic organizations that are capable of evolution can only be based on faith, a notoriously dangerous route to scientific progress."
"Kauffman takes it for granted that if it is possible to write down on paper a closed peptide cycle and a set of catalyzed ligations leading from monomeric amino acids to the peptides of the cycle, then that cycle would self-organize spontaneously and come to dominate the chemistry of a reaction system. This... is unlikely because peptide molecules do not have the properties that Kauffman assigns to them." "I have also explored a number of alternative systems with different numbers of amino acids or with inputs of random families of short peptides, and I find that they all encounter similar or more severe difficulties."
"Kauffman assumes that, in sufficiently concentrated solution, the naturally occurring amino acids or some subset of them would condense spontaneously to form a mixture of long peptides in substantial yield. In practice, this would not happen." "The catalytic properties of enzymes are remarkable. They not only accelerate reaction rates by many orders of magnitude, but they also discriminate between potential substrates that differ very slightly in structure. Would one expect similar discrimination in the catalytic potential of peptides of length ten or less? The answer is clearly 'no', and it is this conclusion that ultimately undermines the peptide cycle theory."
"Protein catalysis is dependent on the stable three-dimensional structures of enzymes and enzyme-substrate complexes. Highly specific catalytic activity could only be expected from short peptides if they, too, adopted stable structures." "In fact, short peptides rarely form stable structures, and when they do, the structures are only marginally stable. The synthesis of a decapeptide that would catalyze the ligation in the correct order of two particular pentapeptides out of a mixture of ten pentapeptides that are required to form the five cycle components, while failing to bring about any of the other possible ligations, would present an extremely difficult challenge to peptide chemistry. It seems certain that the additional requirement that this peptide should also catalyze specifically many of the reactions leading from amino acids to the pentamer precursors of the decamers of the cycle could never be met. Of course, the decamers need not be formed only from pairs of pentamers, but the difficulties are no less severe for more complex synthetic networks. There are a number of possible ways in which this difficulty might be circumvented, but none seems relevant to the origin of life." "It is unlikely, therefore, that Kauffman's theory describes any system relevant to the origin of life."
"It is essential to subject metabolist proposals to the same kind of detailed examination and criticism that has rightly been applied to genetic theories." "Because little experimental work has been attempted, appraisal must be based on chemical plausibility." "The lack of a supporting background in chemistry is even more evident in proposals that metabolic cycles can evolve to 'life-like' complexity. The most serious challenge to proponents of metabolic cycle theories--the problems presented by the lack of specificity of most nonenzymatic catalysts--has, in general, not been appreciated. If it has, it has been ignored."
"Theories of the origin of life based on metabolic cycles cannot be justified by the inadequacy of competing theories: they must stand on their own." "Experimental proof that such cycles are stable against the challenge of side reactions is even more important." "The prebiotic syntheses that have been investigated experimentally almost always lead to the formation of complex mixtures. Proposed polymer replication schemes are unlikely to succeed except with reasonably pure input monomers. No solution of the origin-of-life problem will be possible until the gap between the two kinds of chemistry is closed." "Solutions offered by supporters of geneticist or metabolist scenarios that are dependent on 'if pigs could fly' hypothetical chemistry are unlikely to help."
Orgel, Leslie E. January 2008. The Implausibility of Metabolic Cycles on the Prebiotic Earth. Public Library of Science (PLoS) Biology, Vol. 6, No. 1, e18, pp. 5-13.
Leslie E. Orgel, Ph.D. Oxford, was a biochemist who studied life on primitive Earth. He conducted research at Cambridge, the University of Chicago, the California Institute of Technology, and later joined the Chemical Evolution Laboratory of the Salk Institute for Biological Studies in San Diego, California. He died at age 80 in October 2007. The above article was published posthumously.
After the "tree of life"
In a paper about bacteria, two evolutionary biologists write, "we cannot rely exclusively on traditional genealogical relationships." "A single taxonomy will be likely to provide an overly coarse picture". It should be replaced by "more taxonomies based on real biological processes". "Discarding all but one of these process-based taxonomies would be comparable to reducing a person's identity to a single aspect of his or her life, even though he or she might have an effective role in many organizations: professional, artistic, sports, family and so on. To avoid overlooking any of the natural groups, it seems legitimate to propose - rather than a single taxonomy of microbial species - many taxonomies". "We suggest giving up the unique hierarchy as the reference classification system and instead encourage the production of a comprehensive interactive database in which an individual could possibly belong to overlapping taxonomical groups." "Any organism can then be characterized by many names because it can belong to more than one group at once."Bapteste, Eric, Yan Boucher. 2008. Lateral gene transfer challenges principles of microbial systematics. Trends in Microbiology, Vol. 16, No. 5, pp. 200-207.
Epigenetics (meaning "above" genetics, as in controlling elements)Evolutionists are starting to turn to epigenetics to explain macroevolutionary changes. However, most of them do not know much about molecular biology, and they are scrambling to catch up. "During the twentieth century, evolutionary and molecular biology diverged". "Few scientists were trained in both fields, and many biology departments were split up." "Most evolutionary biologists emphasize... variation". "Inferences about the historical mechanisms that generate variation are usually drawn from patterns of association". "The weakness is that statistical associations are not reliable indicators of causality." "Claims that are based solely on associations remain standard in the field." "Evolutionary biologists will need to be trained in molecular biology".-- Dean, Antony M., Joseph W. Thornton. September 2007. Mechanistic approaches to the study of evolution: the functional synthesis. Nature Reviews Genetics, Vol. 8, pp. 675-688.
Here is some of what real molecular biologists have learned, beginning with a March 2008 report in Science News magazine: "Many people regard ribonucleic acid, as RNA is formally known, as 'just a middleman between DNA and protein,' says Claes Wahlestedt, a neuroscientist and genome researcher at the Scripps Research Institute in Jupiter, Fla. Shuttling genetic information from DNA to a cell's protein factories has long been recognized as RNA's day job, summarized" as "DNA makes RNA makes protein." "Some researchers estimate that as much as 98 percent of the human genome is copied into RNA, says Sofie Salama of the University of California, Santa Cruz." "Initial observations of the genome showed islands of protein-coding genes separated by vast oceans of DNA--sometimes called junk DNA--where nothing happened. That would mean that only about 2 percent of the human genome is transcribed into RNA. But recent efforts to map all of the RNA transcripts show that virtually every base pair of DNA in the human genome is copied into at least one RNA molecule."
"More than 20 classes of noncoding RNA have been discovered in the past decade. Many of these RNAs are much smaller than their protein-coding cousins, the messenger RNAs. Some noncoding RNAs contain a mere 20 nucleotides, the chemical units corresponding to letters in the genetic alphabet. Scientists used to throw away such short bits of RNA, thinking the tiny pieces were nothing more than breakdown products of larger molecules--basically garbage, Wahlestedt says."
"Researchers now know that noncoding RNAs get involved in virtually everything that happens in or to a cell, says Georges St. Laurent III, a computational and molecular biologist at George Washington University in Washington, D.C." "They monitor temperature, chemical conditions, electrical currents, and other signals from the environment and then tell the cell how to respond."
"One class of noncoding RNAs, known as microRNAs, modulates production of proteins. MicroRNAs get their name from their minuscule size--most are only about 22 nucleotides long. These short pieces of RNA find and bind to complementary sequences in messenger RNAs. Usually that binding causes the ribosome, the protein-building machinery in a cell, to grind to a halt. The ribosome remains paused until other signals allow it to resume making protein or until the RNA message is destroyed." " 'It's not only important that you make a particular protein, but when and where you make it,' Salama says."-- Tina Hesman Saey. March 1, 2008. Micromanagers: New classes of RNAs emerge as key players in the brain. Science News, Vol. 173, No. 9, pp. 136-137.
Non-coding RNAs have risen from "junk" to "drivers of complexity".42 "Sequencing the genomes of 85 species has revealed that in any given organism, increasing biological complexity is correlated with an increasing number of non-protein-coding DNA sequences and not, as previously assumed, with an increasing number of protein-coding genes."42 "The sheer number of non-coding RNAs is estimated in the 100s of thousands."15 "It is clear that tens of thousands may operate within a cell".42
"Interference and activation can be caused by the same transcript".4 "A large part of the transcriptional activity in the human genome is derived from repeat sequences".42 "Repeat elements... occupy 40-45% of a typical mammalian genome".4 "Alu repetitive elements constitute 10% of the human genome".31 "Repeat elements, such as the Alu family in humans and B2 in mice have provided regulatory signals for RNA PolII transcription."7 "Some of the Alu elements... may have functions in stress response, chromatin organization or signaling events in the early embryo. Alu transcripts are... activated by heat shock and DNA damaging agents".42
There are levels of non-coding RNA regulation that have yet to be discovered.42 Studying the old "junk" transcripts can lead to understanding hidden layers of cell regulation and how deregulation can lead to the understanding of human disease.42 "The scientific community is getting more aware of the importance of the formerly abandoned 'junk' DNA. What we have learned so far is likely just the tip of the iceberg."42
It is clear that biological complexity depends less on gene number and more on how those genes are used. Researchers are realizing that regulation is on multiple levels32; there are intricate feedback loops.6 Stretches of DNA can be inactivated by attaching methyl groups. Tiny embryos need to grow according to a body plan organized in steps that have to happen at the right time in the right sequence. Their cells use timers and spatial signals to guide their growth. For example, a signal chemical is made at one end of an embryo and spreads out. Cells act according to how much signal chemical reaches them. Signal chemicals spreading from opposite ends of an embryo can interact to coordinate construction.28
In small genomes, such as yeast, the parts of DNA that regulate a gene are next to the gene. In more complex genomes, such as human and mouse, they can be far apart. Cells have ways, still unknown, of moving sections of chromosomes next to each other to get the right parts together to control gene expression.11 This happens constantly.
To respond to a rapidly changing environment, a creature's genes have to be turned on and off in a highly coordinated way. The genetic network must be stable under a broad range of conditions, but flexible enough to recognize and respond to important signals when things around it change. This operating at the brink of order and chaos is well known to systems scientists. They call such systems critical. This property has now been recognized in plants, animals, and microbes. It allows them to quickly detect and respond to external stimuli, small or large.4
In another surprise to evolutionists, genes they have long called vestigial junk have a clear purpose. Some genes are not transcribed into proteins, yet they seem related to working genes. So evolutionists figured these were leftover copying mistakes, called them pseudogenes (fake genes), and ignored them. Researchers studying cancer finally took a look at a pseudogene in action, and found that it competed with its working gene for the same non-coding regulatory RNA. Thus the gene and pseudogene act as decoys for one another, and affect the regulation of other transcripts. The pseudogene "is not a non-functional relic, but a modulator of gene expression." This "could have major implications for understanding mechanisms of disease, and of cancer in particular." "The authors find similar associations between other well-known cancer-associated genes and their corresponding pseudogenes."
Poliseno, Laura, Leonardo Salmena, Jiangwen Zhang, Brett Carver, William J. Haveman, Pier Paolo Pandolfi. 24 June 2010. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature, Vol. 465, pp. 1033-1038.
Rigoutsos, Isidore, Frank Furnari. 24 June 2010. Decoy for microRNAs. News & Views, Nature, Vol. 465, pp. 1016-1017.
The Mind of the Evolutionist
" 'Contemporary evolutionary thinking maintains that smaller island mammals will rapidly grow larger towards the optimal size, while bigger animals will rapidly shrink.' " Evolutionists call this Optimal Body Size Theory, or OST. A member of a research team was interviewed about their study that "looked at a theoretical optimum body size towards which mammals are expected to grow, on both island communities and on the mainland." " 'There is a tendency to believe that big animals become very small on islands, and small animals become very big, due to limited resources or lack of competition. I've shown that this is just not true, at least not as a general rule.' " "Incorporating large data sets that compared body sizes on various islands and on mainland communities, Dr. Meiri and his colleagues found no such tendency for bizarrely-sized animals to develop on islands. 'We concluded that the evolution of body sizes is as random with respect to "isolation" as on the rest of the planet. This means that you can expect to find the same sort of patterns on islands and on the mainland.' Dr. Meiri attributes our widely held misperceptions about 'dragons and dwarfs' to the fact that people tend to notice the extremes more if they are found on islands." "Darwin's fascination with the Galapagos island chain... is just one example." " 'I think it's purely a psychological bias,' Dr. Meiri concludes. 'It's just magical thinking. Nothing more.' "-- 'Magical Thinking' About Islands an Illusion? Biologist Refutes Conventional Thinking on Evolution. July 8, 2010. Science Daily, online news release.
"We found no support for any of the predictions of the optimal size theory." "The concept of a single optimal body size is not supported by the data that were thought most likely to show it." "It is remarkable that this theory fails to apply under the circumstances which best match its predictions (on islands)."-- Raia, Pasquale, Francesco Carotenuto, Shai Meiri. 2010. One size does not fit all: no evidence for an optimal body size on islands. Global Ecology and Biogeography, Vol. 19, pp. 475-484.
Evolutionists claim to rely only on natural forces, but natural forces cannot design and build new plants and animals. So they add magical thinking, the perfect description of the evolutionist mind.
Zombie science
"Although the classical ideal is that scientific theories are evaluated by a careful teasing-out of their internal logic and external implications, and checking whether these deductions and predictions are in-line-with old and new observations; the fact that so many vague, dumb or incoherent scientific theories are apparently believed by so many scientists for so many years is suggestive that this ideal does not necessarily reflect real world practice. In the real world it looks more like most scientists are quite willing to pursue wrong ideas for so long as they are rewarded with a better chance of achieving more grants, publications and status."
To say "that the theory is phoney, and always was phoney, and this is why it so singularly fails to predict reality is regarded as simplistic, crass, merely a sign of lack of sophistication. And anyway, there are... the reputations of numerous scientists who are now successful and powerful on the back of the phoney theory, and who by now control the peer review process (including allocation of grants, publications and jobs) so there is a powerful disincentive against upsetting the apple cart."
"Zombie science is science that is dead but will not lie down." "Zombie science is supported because it is useful propaganda. Zombie science is deployed in arenas such as political rhetoric, public administration, management, public relations, marketing and the mass media generally. It persuades, it constructs taboos, it buttresses some kind of rhetorical attempt to shape mass opinion. Indeed, zombie science often comes across in the mass media as being more plausible than real science."Charlton, Bruce G. 2008. Zombie science: A sinister consequence of evaluating scientific theories purely on the basis of enlightened self-interest. Medical Hypotheses, Vol. 71, pp. 327-329.
|
Darwin
is liked by evolutionists because he liberated science
from the straitjacket of observation and opened the
door to storytellers. This gave professional evolutionists
job security so they can wander through biology labs
as if they belong there.
---
David Coppedge
Speaking of Science, Creation Matters, May/June 2003 |

**********
1. Alberts, Bruce, Alexander Johnson, Julian Lewis, Martin Raff,
Keith Roberts, Peter Walter. 2008. Molecular Biology of The Cell,
5th edition. Garland Science, New York.2. Anderson, G. M. 1996. Thermodynamics of Natural Systems. John Wiley & Sons, Toronto.
3. Baker, Monya. 10 February 2011. Genomes in Three Dimensions. Nature, Vol. 470, pp. 289-294.
4. Balleza, Enrique, Elena R. Alvarez-Buylla, Alvaro Chaos, Stuart Kauffman, Ilya Shmulevich, Maximino Aldana. June 2008. Critical Dynamics in Genetic Regulatory Networks: Examples from Four Kingdoms. PLoS ONE, Vol. 3, No. 6, pp. 1-10.
5. Bergman, Jerry. August 2000. Do any vestigial organs exist in humans? Technical Journal, Vol. 14, No. 2, pp. 95-98.
6. Brandman, Onn, Tobias Meyer. 17 October 2008. Feedback Loops Shape Cellular Signals in Space and Time. Science, Vol. 322, No. 5900, pp. 390-395.
7. Carninci, Piero, Jun Yasuda, Yoshihide Hayashizaki. 2008. Multifaceted mammalian transcriptome. Current opinion in Cell Biology, Vol. 20, pp. 274-280.
8. Cemic, Ladislav. 2005. Thermodynamics in Mineral Sciences. Springer, New York.
9. Coates, Michael I. 26 March 2009. Beyond the Age of Fishes. Nature, Vol. 458. pp. 413-414.
10. Dagan T., W. Martin. 2006. The tree of one percent. Genome Biology, Vol. 7, No. 10, pp. 118.
11. Dekker, Job. 28 March 2008. Gene Regulation in the Third Dimension. Science, Vol. 319, pp. 1793-1794.
12. Doolittle, W. Ford, Eric Bapteste. February 13, 2007. Pattern pluralism and the Tree of Life hypothesis. Proceedings of the National Academy of Sciences of the United States of America, Vol. 104, No. 7, pp. 2043-2049.
13. Doolittle, W. Ford. 25 June 1999. Phylogenetic Classification and the Universal Tree. Science, Vol. 284, No. 5423, pp. 2124-2128.
14. Duke University Medical Center. "Evolution Of The Human Appendix: A Biological 'Remnant' No More." ScienceDaily 21 August 2009. Retrieved 12 September 2009 <http://www.sciencedaily.com /releases/2009/08/090820175901.htm>.
15. Fahraeus, Robin, Marc Blondel. 2008. Editorial: RNA-assisted protein folding. Biotechnology Journal, Vol. 3, pp. 967-969.
16. Franze, Kristian, Jens Grosche, Serguei N. Skatchkov, Stefan Schinkinger, Christian Foja, Detlev Schild, Ortrud Uckermann, Kort Travis, Andreas Reichenbach, Jochen Guck. May 15, 2007. Muller cells are living optical fibers in the vertebrate retina. PNAS Vol. 104, No. 20, pp. 8287-8292. DOI: 10.1073/pnas.0611180104
17. Gillooly, James F., Andrew P. Allen, Geoffrey B. West, James H. Brown. January 4, 2005. The rate of DNA evolution: Effects of body size and temperature on the molecular clock. Proceedings of the National Academy of Sciences of the United States of America, Vol. 102, No. 1, pp. 140-145.
18. Gish, Duane T., PhD. Biochemistry. January 2007. A Few Reasons an Evolutionary Origin of Life Is Impossible. Impact #403, Acts and Facts, Institute for Creation Research.
19. Glass, John I., Nacyra Assad-Garcia, Nina Alperovich, Shibu Yooseph, Matthew R. Lewis, Mahir Maruf, Clyde A. Hutchison III, Hamilton O. Smith, J. Craig Venter. January 10, 2006. Essential genes of a minimal bacterium. Proceedings of the National Academy of Sciences of the United States of America, Vol. 103, No. 2, pp. 425-430.
20. Jacob, Francois. June 10 1977. Evolution and Tinkering. Science, New Series, Vol. 196, Issue 4295, pp. 1161-1166.
21. Khalturin K, Anton-Erxleben F, Sassmann S, Wittlieb J, Hemmrich G, et al. 2008. A novel gene family controls species-specific morphological traits in Hydra. PLoS Biol 6(11): e278. doi:10.1371/journal.pbio.0060278
22. Koonin, Eugene V. 12 February 2009. Darwinian evolution in the light of genomics. Nucleic Acids Research, Vol. 37, No. 4, pp. 1011-1034.
23. Koonin, Eugene V. 20 August 2007. The Biological Big Bang model for the major transitions in evolution. Biology Direct, Vol. 2:21, pp. 1-17.
24. Labin, A.M., E. N. Ribak. 16 April 2010. Retinal Glial Cells Enhance Human Vision Acuity. Physical Review Letters, Vol. 104, No. 15, pp. 158102-1 to 4. DOI: 10.1103/PhysRevLett.104.158102
25. Labin, Amichai M., Shadi K. Safuri, Erez N. Ribak, Ido Perlman. 8 July 2014. Muller cells separate between wavelengths to improve day vision with minimal effect upon night vision. Nature Communications, Vol. 5, No. 4319, pp. 1-9. DOI: 10.1038/ncomms5319
26. Lawton, Graham. 21 January 2009. Why Darwin was wrong about the tree of life. New Scientist Magazine, issue 2692.
27. Lee, Michael S.Y., James B. Jago, Diego C. Garcia-Bellido, Gregory D. Edgecombe, James G. Gehling, John R. Paterson. 30 June 2011. Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature, Vol. 474, pp. 631-634.
28. Lewis Julian. 17 October 2008. From Signals to Patterns: Space, Time, and Mathematics in Developmental Biology. Science, Vol. 322, pp. 399-403.
29. Lowe, Craig B., Gill Bejerano, David Haussler. May 8, 2007. Thousands of human mobile element fragments undergo strong purifying selection near developmental genes. Proceedings of the National Academy of Sciences of the United States of America, Vol. 104, No. 19, pp. 8005-8010.
30. Lunyak, Victoria V., Gratien G. Prefontaine, Esperanza Nunez, Thorsten Cramer, Bong-Gun Ju, Kenneth A. Ohgi, Kasey Hutt, Rosa Roy, Angel Garcia-Diaz, Xiaoyan Zhu, Yun Yung, Lluis Montoliu, Christopher K. Glass, Michael G. Rosenfeld. July 13, 2007. Developmentally Regulated Activation of a SINE B2 Repeat as a Domain Boundary in Organogenesis. Science, Vol. 317, No. 5835, pp. 248-251.
31. Makalowski, Wojciech. May 23, 2003. Not Junk After All. Science, Vol. 300, No. 5623, pp. 1246-1247.
32. Makeyev, Eugene V., Tom Maniatis. 28 March 2008. Multilevel Regulation of Gene Expression by MicroRNAs. Science, Vol. 319, pp. 1789-1790.
34. Pace, John K. II, Clement Gilbert, Marlena S. Clark, Cedric Feschotte. November 4, 2008. Repeated horizontal transfer of a DNA transposon in mammals and other tetrapods. Proceedings of the National Academy of Sciences, Vol. 105, No. 44, pp. 17023-17028.
35. Pilcher, Helen. 19 January 2013. Genes from nowhere: Orphans with a surprising story. New Scientist, No. 2900, pp. 38-41.
36. Prigogine, Ilya, Gregoire Nicolis, Agnes Babloyants. 1972. Thermodynamics of Evolution. Physics Today, Vol. 25, No. 11, pp. 23-44.
37. Reichenbach, Andreas, Andreas Bringmann. May 2013. New functions of Muller cells. Glia, Vol. 61, No. 5, pp. 651-678. DOI: 10.1002/glia.22477
38. Richardson, Michael K., James Hanken, Mayoni L. Gooneratne, Claude Pieau, Albert Raynaud, Lynne Selwood, Glenda M. Wright. July 1997. There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development. Anatomy and Embryology, Vol. 196, No. 2, pp. 91-106.
39. Shu, D.-G., S. Conway Morris, J. Han, Z.-F. Zhang, K. Yasui, P. Janvier, L. Chen, X.-L. Zhang, J.-N. Liu, Y. Li, H.-Q. Liu. 30 January 2003. Head and backbone of the Early Cambrian vertebrate Haikouichthys. Nature, Vol. 421, pp. 526-529.
40. Smith, H.F., R.E. Fisher, M.L. Everett, A.D. Thomas, R. Randal Bollinger, W. Parker. October 2009. Comparative anatomy and phylogenetic distribution of the mammalian cecal appendix. Journal of Evolutionary Biology, Vol. 22, No. 10, pp. 1984-1999. doi:10.1111/j.1420-9101.2009.01809.x
41. Smith, Heather F., William Parker, Sanet H. Kotze, Michel Laurin. 2013. Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors. Comptes Rendus Palevol, 16 pages. doi:10.1016/j.crpv.2012.12.001
42. Szell, Marta, Zsuzsanna Bata-Csorgo, Lajos Kemeny. 2008. The enigmatic world of mRNA-like ncRNAs: Their role in human evolution and in human diseases. Seminars in Cancer Biology, Vol. 18, pp. 141-148.
43. van Steensel, Bas, Job Dekker. October 2010. Genomics tools for unraveling chromosome architecture. Nature Biotechnology, Vol. 28, No. 10, pp. 1089-1093.
44. Weaver, Robert F. 2008. Molecular Biology, Fourth Edition. McGraw-Hill, New York. pp. 660-680.
45. Wissler, Lothar, Jurgen Gadau, Daniel F. Simola, Martin Helmkampf, Erich Bornberg-Bauer. January 24, 2013. Mechanisms and dynamics of orphan gene emergence in insect genomes. Genome Biology and Evolution Advance Access. doi:10.1093/gbe/evt009
46. Wood, Richard D., Michael Mitchell, John Sgouros, Tomas Lindahl. 16 February 2001. Human DNA Repair Genes. Science, Vol. 291, No. 5507, pp. 1284-1289.
47. Wu D-D, Irwin DM, Zhang Y-P. 2011. De Novo Origin of Human Protein-Coding Genes. PLoS Genet 7(11): e1002379. doi:10.1371/journal.pgen.1002379
48. Zhu, Min, Wenjin Zhao, Liantao Jia, Jing Lu, Tuo Qiao, Qingming Qu. 26 March 2009. The oldest articulated osteichthyan reveals mosaic gnathostome characters. Nature, Vol. 458, pp. 469-474.
2006-2015
John Michael Fischer
mike@newgeology.us
John Michael Fischer
mike@newgeology.us
from crev.info
No comments:
Post a Comment